近几十年, 人类活动使陆地生态系统的固氮量成倍增加[1 -2 ] , 目前热带、亚热带地区的氮肥使用量占全球使用量的 40%[3 ] , 据预测, 到 2020 年, 全球约2/3的氮输入将发生在热带、亚热带地区[1 ] 。我国亚热带地区也存在高氮沉降量现象, 如太湖的常熟生态站大气氮湿沉降量在 2001—2003 年间约为 27 kg/(hm2 ·a)[4 ] , 广东鼎湖山自然保护区在 20 世纪90年代末的降水氮沉降为 38.4 kg/(hm2 ·a)[5 ] 。氮沉降的不断增加对我国森林生态系统的结构和功能造成多方面的影响[6 ] 。
乔木生长对于维持森林生态系统功能十分重要, 以往研究表明, 氮沉降的加剧对森林生长和生态系统功能存在不同影响。一方面, 由于陆地生态系统多存在氮限制, 短期内有效氮的增加会促进森林生长, 加速生物量的积累, 从而提高陆地生态系统的固碳量[7 -8 ] , 同时氮添加对树木生长的促进程度还受树龄及不同树种氮利用方式等因素的影 响[9 -10 ] ; 另一方面, 长期施肥实验表明, 过量氮的输入会引起生态系统氮饱和, 导致一系列环境负效应, 如养分失衡、土壤酸化、盐基阳离子流失、有害离子富集和植物抗性下降等。超过生态系统氮负荷之后, 氮添加会抑制植物的生长[11 -12 ] , 引起生态系统衰退[13 -14 ] 。国内外研究发现, 温带森林多为氮限制生态系统, 氮沉降可在一定程度上促进森林树木的生长[15 -17 ] 。与此同时, 有关氮沉降对热带森林的施肥效果仍存在一定争议, 相对于氮元素, 通常认为热带地区磷对生长的限制作用更强, 一些研究也证实氮沉降对森林生长没有影响[18 -20 ] 。然而LeBauer等[21 ] 对 16 个热带森林研究后发现, 部分热带森林类型表现出明显的氮限制特点。由于森林生长对氮沉降的响应存在诸多不确定性[22 -24 ] , 因此对武夷山地区天然米槠林进行氮添加背景下的生长研究有利于评估氮沉降对该区域亚热带常绿阔叶林生态系统的影响。
本研究分别用手工测量方法(传统方法)及生长测量仪对武夷山样地内米槠(Castanopsis carlesii )进行一年的连续监测, 主要目的是研究: 1) 将米槠全年进行生长阶段划分后, 不同生长阶段米槠的径向生长对氮添加的响应; 2) 不同径级米槠的生长对氮添加的响应是否存在差异。
1 研究区概况
研究区位于福建武夷山国家自然保护区, 地处武夷山脉北端, 武夷山市、建阳市、光泽县和邵武市四县(市)交界处, 地理范围为 117°27′—51′E, 27°33′—54′N。该区属典型的亚热带季风气候, 夏季高温多雨, 冬季温和湿润。年均气温 8.5~18°C, 年均相对湿度 78%~84%, 年均降水量 1486~2150 mm, 无霜期 253~273 天, 年平均雾日达 120 天[25 ] 。研究区内地带性植被为亚热常绿阔叶林, 植被类型属于保存比较完好的米槠群落, 物种丰富, 层次分明[26 ] , 地带性土壤为红黄壤[25 ] 。
林内闭郁度为 0.9, 总体平均林分密度(SD)为2613±22 株/hm2 , 米槠平均胸径(SD)为 24.5±8.0 cm,平均树高 19.6±4.8 m (表 1 ), 群落层次分明, 米槠为样地所在群落优势种。其他主要物种组成: 泡花润楠(Machilus pingii )、树参(Dendropanax dentiger )、丝栗栲(Castanopsis fargesii )、拟赤杨(Alniphyllum fortunei )、虎皮楠(Daphniphyllum oldhamii )、弯蒴杜鹃(Rhododendron henryi )、柃木(Eurya japonica )。
2 样点布设和研究方法
2.1 样点布设及实验方法
实验样地位于 117°44′52.91″E, 27°42′31.06″N, 海拔629 m, 平均坡度为35°, 坡向为南偏西13°。共设12块样地, 样方面积为20 m×20 m, 并设置对照(0)、低氮(50 kgN/(hm2 ·a))、中氮(100 kgN/ (hm2 ·a))和高氮(150 kg N/(hm2 ·a))共4个氮添加水平的处理, 每个处理重复3次。自2011年起, 每月在固定时间对样地施加硝酸铵(NH4 NO3 ), 施肥时间为每年的 3—11 月。根据处理水平的要求, 将每个样方每次所需喷施的氮肥溶解在 30 L 水中后, 用背式喷雾器在各样地人工来回均匀喷洒, 对照样地喷施同量的水。
对样方内所有胸径(diameter at breath height, DBH) ≥5 cm的米槠于1.3 m高处安装自制生长环, 并测定其初始胸径值。2014年1月至12月, 每月的月末用电子游标卡尺测量其胸围变化量。同时, 在每种处理水平下选取1棵干形较直、胸径及树高均接近样地内同物种平均值的米槠作为标准树(表2 ), 安装自动胸径记录仪(Dendrometer-DRL26, 捷克), 每隔 30 分钟记录 1 次数据, 对采集的数据用USB 红外转输下载。树木径向生长测量仪是一种可以自动记录树木原位生长精确数据的电子仪器, 与传统的监测方法(直接测量胸径)相比, 该仪器能够识别树干水分引起的日际收缩和膨胀波动[27 ] 。本研究共安置 4 个带状记录仪, 自动记录生长数据。采用的树木生长数据来源于自制生长环法和自动记录仪法两部分, 时间范围为 2014 年 1—12月。
2.2 数据分析
2.2.1 判断生长时期
依据 Deslauriers 等[28 ] 的方法, 结合实验样地的经纬度及海拔, 树木的生长时期由连续 15 日的径向日变化均值确定, 该方法比直接观察胸径值变化趋势更准确。2014 年 3 月 15 日至 2015 年 3 月 1日, 使用单样本t 检验对每连续15日的径向日变化均值(即连续15日的每日径向变化值取平均)与0做比较, 当值的分布显著大于零时, 认为开始发生明显的径向生长[29 ] 。树木的径向日变化量由Bouriaud等[30 ] 提出的最大值法计算而来, 从记录米槠茎干每日生长的 48 个原始数据中提取日最大值, 计算连续两日的最大值之差, 求得树木茎干的日变化量。
2.2.2 生长速率
根据检测得到的每月胸围生长量以及树木初始胸径值, 计算得到每月的胸径值, 进而计算出胸径相对生长率。每月胸径相对生长率(%)=(本月胸径值-上月胸径值)/上月胸径值×100%, 全年胸径相对生长率(%)=12 个月的胸径增长量/初始胸径值× 100%。将所有样方内的米槠按照初始胸径的大小分为3个径级(5~15 cm, 15~25 cm 和>25 cm), 以确保每个径级中树木数量接近, 并分别计算各个径级的相对生长率。
本文统计分析使用 SPSS19.0, 图形绘制使用Origin8.6, 单因素方差分析采用 Duncan 多重检验法进行两两比较, 检验不同氮添加处理差异的显著性, 在图中用不同字母表示处理间差异性。
3 结果
3.1 米槠季节生长变化规律
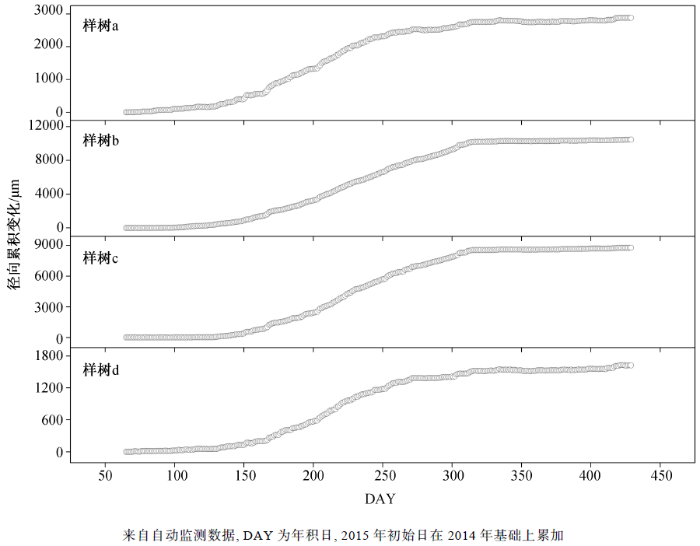
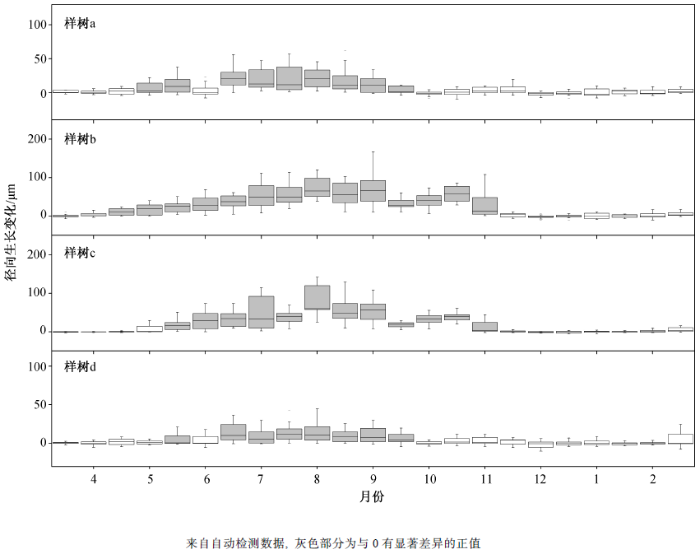
图 1显示米槠全年的茎干累计生长变化动态, 可以看到4棵样树的全年径向生长表现出一致的变化规律, 冬季及初春茎干增长极为缓慢, 夏季表现为快速的持续增长, 随后生长速率立刻放缓。使用Deslauriers 等[28 ] 判断树木不同生长阶段的方法, 对每连续 15 天的日径向变化值进行分析, 结果如图2 所示。对4棵样树分别进行分析, 发现第一个与 0 有显著差异的正值均出现在4月中旬至5 月中旬。此阶段为茎干水分恢复阶段(5 月), 树木茎干表现为因吸水而出现的可逆性逐渐增粗。之后, 样树 a和 d 在 6 月初出现一个与 0 无显著差异的值, 此阶段为水分恢复与茎干生长之间的平台期。在 10—11月, 4棵样树先后出现最后一个与0有显著差异的正值。因此, 可将 6—10 月划分为茎干快速生长阶段, 树木茎干在此阶段快速增粗, 径向增长加快。11—4 月则为茎干缓慢生长阶段, 此阶段没有出现与0有显著差异的正值, 米槠茎干径向生长速率大幅放缓。
图1 2014—2015年米槠全年径向累计变化
Fig. 1 Cumulative radial variation of Castanopsis carlesii from 2014 to 2015
图2 2014 年3月15日—2015年3月1日, 以15日为1组的米槠径向变化分布
Fig. 2 Stem radius variation distribution of Castanopsis carlesii grouped at 15 days from 15 Mar. 2014 to 1 Mar. 2015
3.2 氮添加对米槠胸径生长速率的影响
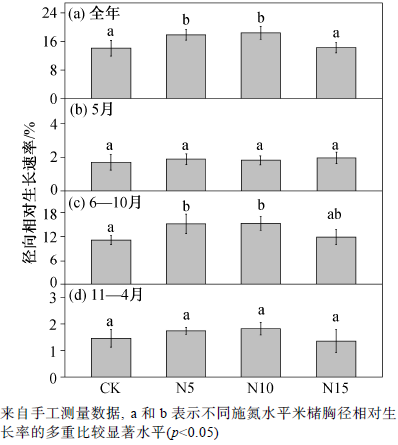
在 2011 年施氮处理前, 对照、低氮、中氮和高氮处理样地的米槠平均胸径(SD)为 22.4±6.2, 19.1±5.7, 20.1±7.6和20.1±6.8 cm, 各处理间胸径值差异不显著。施氮处理后, 低氮和中氮处理均显著提高了米槠的全年径向生长速率(p <0.05), 高氮处理下米槠胸径生长速率与对照无显著差异, 低氮和中氮处理分别使米槠胸径相对生长率比对照增加26.4%和30.3% (图 3 (a))。
通过将米槠的不同生长时期分别比较后发现, 在水分恢复期(5 月), 氮添加对米槠胸径生长速率无显著影响。胸径增长速率分别为对照 1.7%, 低氮处理 1.9%, 中氮处理1.8%, 高氮处理 2.0% (图3 (b))。在米槠的快速生长期(6—10 月), 不同处理下胸径生长速率与全年表现出一致趋势, 低氮和中氮处理对米槠生长有显著促进作用(p <0.05), 分别比对照增加 35.7%和 37.1%, 高氮处理使径向相对生长率比对照增加 6.3%, 差异不显著(图 3 (c))。在缓慢生长阶段(11—4 月), 各处理间均无显著差异(图3 (d))。
图3 不同氮浓度处理米槠径向相对生长速率变化
Fig. 3 Responses of Castanopsis carlesii diameter at breast height (DBH) relative growth rate under different N treatments
3.3 不同径级米槠胸径生长率对氮添加的响应
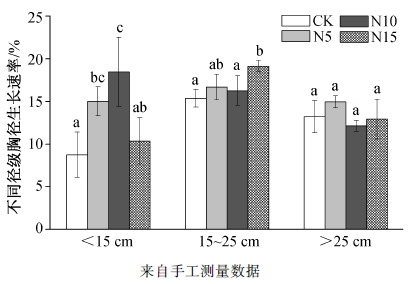
分析表明, 不同径级米槠胸径相对生长率对氮添加的响应存在差异。如图 4 所示, 在5~15 cm径级, 低氮和中氮处理对胸径生长速率有显著促进作用(p <0.05), 其中, 中氮处理对生长的促进作用最显著。与对照样地相比, 低氮和中氮处理 DBH 分别增加 71.7%和 111.0%, 高氮处理影响不显著; 在15~25 cm 径级内, 高氮处理显著提高了米槠胸径相对生长率(24.5%, p <0.05), 低氮和中氮处理虽表现为促进作用, 但不显著; 对于 25 cm 径级以上的米槠来说, 胸径相对生长率在各处理间均无显著差异。
图4 不同径级米槠全年径向相对生长速率对氮添加的响应
Fig. 4 Responses of Castanopsis carlesii DBH relative growth rate under different N treatments among different tree diameter classes
4 讨论
4.1 武夷山米槠生长阶段的判定
选择合适的时间尺度对自动记录数据进行分析, 可以更准确地识别树木的生长阶段。Deslau-riers 等[28 ] 认为, 在较低海拔地区, 选择 10 日以上的时间尺度, 较易识别树木茎干生长的主要阶段。根据样地所处的位置以及海拔, 本研究选择 15 日的时间尺度以及 4 株样树来识别米槠全年的不同生长阶段。结果表明, 样树a和d的径向生长在6月初至6月中旬有一段时间表现为非明显增长的平台期(图 2 ), 以此来区分树木在本年内的快速生长阶段。样树b和c未识别出这一时期, 可能主要是因为亚热带地区春季降雨量大且持续时间较长, 树木的补水阶段与夏季的径向快速生长阶段实现快速过渡。
4.2 氮添加对生长速率的影响
氮添加在总体水平上增加了米槠的全年胸径相对生长速率, 说明研究区内氮素是米槠生长的限制因素。研究区内米槠在不同氮添加水平下的响应不同, 除高氮处理外, 低氮和中氮处理均显著提高了米槠的全年径向生长速率。这与我国目前在热带及亚热带森林生态系统的多数氮沉降试验结果一致, 氮沉降会促进受氮限制的年轻森林中树木的生 长[22 ,31 -33 ] , 树木对氮沉降的响应也受树木种类、研究地点和施肥年限等因素的影响。樊后保等[31 ] 对亚热带杉木的生长研究结果表明, 经过 3 年的施氮处理后, 中氮处理(120 kg N/(hm2 ·a))和高氮处理(240 kg N/(hm2 ·a))均明显促进杉木胸径的生长。在鼎湖山的研究也发现, 中氮处理(100 kg N/(hm2 ·a))使马尾松林胸径生长增加 77.8%[32 ] , 高氮处理则将逐渐对乔木幼苗生长产生负面效应[22 ] 。通过对米槠的季节性生长分析发现, 氮添加的生长促进作用主要在水热条件较好的湿润季节产生, 这可能是由于研究地区夏季高温多雨, 土壤氮的周转速率增强, 提高了植物生长过程中氮素的可利用性[34 ] , 当氮素为限制因子时, 额外的氮输入可通过增加土壤和凋落物有效氮[35 ] 、改变基质的 C/N 比[6 ] 及改变土壤微生物组成[36 ] 等方式, 短期提高土壤的氮矿化速率, 进一步提高土壤氮素的可利用性。
以往的研究表明, 不同的树木径级对氮添加表现出不同的响应。郑威等[37 ] 对湖南樟树人工林的施肥实验表明, 施氮能够明显提高8 cm和 30 cm径阶的林木生长率。本研究样地内物种组成丰富, 米槠作为样地所在群落的优势种, 树木径级的不同可通过影响树木对光竞争、养分竞争及维持呼吸等方面来影响米槠的径向生长[38 -39 ] 。低径级米槠对低氮和中氮处理响应强烈, 可能是由于低径级米槠生长代谢较为活跃, 冠层的快速增长导致对氮素需求较大。中等径级的米槠为样地中的优势个体, 除中氮处理外, 其他氮浓度处理下胸径相对生长率均随胸径增粗而提高, 但由于部分资源要分配给生殖生长以及根系和树干呼吸的增强[40 ] , 高氮处理使其相对生长速率显著提升。随着径级继续增大, 米槠多为老龄个体, 生长活力下降[41 ] , 光合作用产物主要用于维持性呼吸代谢[39 ] , 因而其径向生长对施氮可能无明显响应。
The authors have declared that no competing interests exist.
参考文献
文献选项
[1]
Galloway J N Schlesinger W H Levy H et al. Nitrogen fixation: anthropogenic enhancement-envir-onmental response.
Global Biogeochemical Cycles , 1995 , 9 (2 ): 235 -252
[本文引用: 2]
[2]
Vitousek P M Farrington H. Nutrient limitation and soil development: experimental test of a biogeoche-mical theory.
Biogeochemistry , 1997 , 37 (1 ): 63 -75
[本文引用: 1]
[3]
Matthews E. Nitrogenous fertilizers: global distri-bution of consumption and associated emissions of nitrous oxide and ammonia.
Global Biogeochemical Cycles , 1994 , 8 (4 ): 411 -439
[本文引用: 1]
[4]
王小治 , 朱建国 , 高人 , 等 . 太湖地区氮素湿沉降动态及生态学意义: 以常熟生态站为例
. 应用生态学报 , 2004 , 15 (9 ): 1616 -1620
[本文引用: 1]
[5]
周国逸 , 闫俊华 . 鼎湖山区域大气降水特征和物质元素输入对森林生态系统存在和发育的影响
. 生态学报 , 2001 , 21 (12 ): 2002 -2012
[本文引用: 1]
[6]
李德军 , 莫江明 , 方运霆 , 等 . 氮沉降对森林植物的影响
. 生态学报 , 2003 , 23 (9 ): 1891 -1900
[本文引用: 2]
[7]
Matson P Hall S J. The globalization of nitrogen deposition: consequences for terrestrial ecosystems.
Ambio , 2002 , 31 (2 ): 113 -119
[本文引用: 1]
[8]
Vejre H Callesen I Vesterdal L et al. Carbon and nitrogen in Danish forest soils — contents and distri-bution determined by soil order.
Soilence Society of America Journal , 2003 , 67 (1 ): 335 -343
[本文引用: 1]
[9]
周利勋 , 刘广平 , 王金波 . 落叶松人工林的施肥效应
. 东北林业大学学报 , 2004 , 32 (2 ): 16 -18
[本文引用: 1]
[10]
Hyvönen R Persson T Andersson S et al. Impact of long-term nitrogen addition on carbon stocks in trees and soils in northern Europe.
Biogeochemistry , 2008 , 89 (1 ): 121 -137
[本文引用: 1]
[11]
Thomas R Q Canham C D Weathers K C et al. Increased tree carbon storage in response to nitrogen deposition in the US.
Nature Geoscience , 2010 , 3 (1 ): 13 -17
[本文引用: 1]
[12]
Aber J McDowell W, Nadelhoffer K, et al. Nitrogen saturation in temperate forest ecosystems-Hypotheses revisited.
Bioscience , 1998 , 48 (11 ): 921 -934
[本文引用: 1]
[13]
Neff J C Townsend A R Gleixner G et al. Variable effects of nitrogen additions on the stability and turnover of soil C.
Nature , 2002 , 419 : 915 -917
[本文引用: 1]
[14]
Nordin A Strengbom J Witzell J et al. Nitrogen deposition and the biodiversity of boreal forests: implications for the nitrogen critical load.
Ambio , 2005 , 34 (1 ): 20 -24
[本文引用: 1]
[15]
Solberg S Dobbertin M Reinds G J et al. Analyses of the impact of changes in atmospheric deposition and climate on forest growth in European monitoring plots: a stand growth approach.
Forest Ecology and Management , 2009 , 258 (8 ): 1735 -1750
[本文引用: 1]
[16]
Vries W D Du E Butterbach-Bahl K Short and long-term impacts of nitrogen deposition on carbon sequestration by forest ecosystems
. Current Opinion in Environmental Sustainability , 2014 , 9/10 : 90 -104
[17]
Laubhann D Sterba H Reinds G J et al. The impact of atmospheric deposition and climate on forest growth in European monitoring plots: an individual tree growth model.
Forest Ecology & Management , 2009 , 258 (8 ): 1751 -1761
[本文引用: 1]
[18]
Cusack D F Silver W L Torn M S et al. Effects of nitrogen additions on above- and belowground carbon dynamics in two tropical forests.
Biogeochemistry , 2011 , 104 : 203 -225
[本文引用: 1]
[19]
Mirmanto E Proctor J Green J et al. Effects of nitrogen andphosphorus fertilization in a lowland evergreen rainforest.
Philosophical Transactions of the Royal Society of London Series B — Biological Sciences , 1999 , 354 : 1825 -1829
[20]
Vitousek P M Porder S Houlton B Z et al. Terre-strial phosphorus limitation: mechanisms, implications, and nitrogen-phosphorus interactions.
Ecological Applications , 2010 , 20 (1 ): 5 -15
[本文引用: 1]
[21]
LeBauer D S Treseder K K Nitrogen limitation of net primary productivity in terrestrial ecosystems is globally distributed.
Ecology , 2008 , 89 (2 ): 371 -379
[本文引用: 1]
[22]
李德军 , 莫江明 , 方运霆 , 等 . 模拟氮沉降对南亚热带两种乔木幼苗生物量及其分配的影响
. 植物生态学报 , 2005 , 29 (4 ): 543 -549
[本文引用: 3]
[23]
李德军 , 莫江明 , 方运霆 , 等 . 模拟氮沉降对三种南亚热带树苗生长和光合作用的影响
. 生态学报 , 2004 , 24 (5 ): 876 -882
[24]
吴茜 , 丁佳 , 闫慧 , 等 . 模拟降水变化和土壤施氮对浙江古田山 5 个树种幼苗生长和生物量的影响
. 植物生态学报 , 2011 , 35 (3 ): 256 -267
[本文引用: 1]
[25]
陈仁华 . 武夷山国家级自然保护区土壤的现状与保护
. 林业勘察设计 , 2006 (2 ): 133 -135
[本文引用: 2]
[26]
李明阳 , 吴军 , 时宇 , 等 . 武夷山国家级自然保护区森林土壤涵水量估测及空间分析
. 西南林业大学学报 , 2014 , 34 (1 ): 1 -7
[本文引用: 1]
[27]
Drew D M Downes G M. The use of precision dendrometers in research on daily stem size and wood property variation: a review.
Dendrochronologia , 2009 , 27 (2 ): 159 -172
[本文引用: 1]
[28]
Deslauriers A Rossi S Anfodillo T. Dendrometer and intra-annual tree growth: what kind of information can be inferred.
Dendrochronologia , 2007 , 25 (2 ): 113 -124
[本文引用: 3]
[29]
李兴欢 , 刘瑞鹏 , 毛子军 , 等 . 小兴安岭红松日径向变化及其对气象因子的响应
. 生态学报 , 2014 , 34 (7 ): 1635 -1644
[本文引用: 1]
[30]
Bouriaud O Leban J M Bert D et al. Intra-annual variations in climate influence growth and wood density of Norway spruce.
Tree Physiology , 2005 , 25 (6 ): 651 -660
[本文引用: 1]
[31]
樊后保 , 刘文飞 , 李燕燕 , 等 . 亚热带杉木(Cun-ninghamia lanceolata )人工林生长与土壤养分对氮沉降的响应
. 生态学报 , 2007 , 27 (11 ): 4630 -4642
[本文引用: 2]
[32]
方运霆 , 莫江明 , 周国逸 , 等 . 鼎湖山主要森林类型植物胸径生长对氮沉降增加的初期响应
. 热带亚热带植物学报 , 2005 , 13 (3 ): 198 -204
[本文引用: 1]
[33]
周璋 . 氮磷添加对海南热带山地雨林碳循环的影响[D]
. 北京: 北京大学 , 2013
[本文引用: 1]
[34]
Ouyang X J Zhou G Y Huang Z L et al. Effect of simulated acid rain on potential carbon and nitrogen mineralization in forest soils.
Pedosphere , 2008 , 18 (4 ): 503 -514
[本文引用: 1]
[35]
Gundersen P Emmett B A Kjønaas O J et al. Impact of nitrogen deposition on nitrogen cycling in forests: a synthesis of NITREX data.
Forest Ecology & Management , 1998 , 101 (1/2/3 ): 37 -55
[本文引用: 1]
[36]
Tietema A. Microbial carbon and nitrogen dynamics in coniferous forest floor material collected along a European nitrogen deposition gradient.
Forest Ecology & Management , 1998 , 101 (1 ): 29 -36
[本文引用: 1]
[37]
郑威 , 闫文德 , 梁小翠 , 等 . 氮添加对樟树林生长的影响
. 中南林业科技大学学报 , 2013 , 33 (4 ): 34 -37
[本文引用: 1]
[38]
Kajimoto T Matsuura Y Sofronov M A et al. Above- and belowground biomass and primary productivity of a Larix gmelinii stand near Tura, central Siberia.
Tree Physiology , 1999 , 19 (12 ): 815 -822
[本文引用: 1]
[39]
Murty D Mcmurtrie R E Ryan M G. Declining forest productivity in aging forest stands: a modeling analy-sis of alternative hypotheses.
Tree Physiology , 1996 , 16 (1/2 ): 187 -200
[本文引用: 2]
[40]
Ryan M G Yoder B J. Hydraulic limits to tree height and tree growth.
Bioscience , 1997 , 47 (4 ): 235 -242
[本文引用: 1]
[41]
Hérault B Bachelot B Poorter L et al. Functional traits shape ontogenetic growth trajectories of rain forest tree species.
Journal of Ecology , 2011 , 99 (6 ): 1431 -1440
[本文引用: 1]
Nitrogen fixation: anthropogenic enhancement-envir-onmental response.
2
1995
... 近几十年, 人类活动使陆地生态系统的固氮量成倍增加[1 -2 ] , 目前热带、亚热带地区的氮肥使用量占全球使用量的 40%[3 ] , 据预测, 到 2020 年, 全球约2/3的氮输入将发生在热带、亚热带地区[1 ] .我国亚热带地区也存在高氮沉降量现象, 如太湖的常熟生态站大气氮湿沉降量在 2001—2003 年间约为 27 kg/(hm2 ·a)[4 ] , 广东鼎湖山自然保护区在 20 世纪90年代末的降水氮沉降为 38.4 kg/(hm2 ·a)[5 ] .氮沉降的不断增加对我国森林生态系统的结构和功能造成多方面的影响[6 ] . ...
... [1 ].我国亚热带地区也存在高氮沉降量现象, 如太湖的常熟生态站大气氮湿沉降量在 2001—2003 年间约为 27 kg/(hm2 ·a)[4 ] , 广东鼎湖山自然保护区在 20 世纪90年代末的降水氮沉降为 38.4 kg/(hm2 ·a)[5 ] .氮沉降的不断增加对我国森林生态系统的结构和功能造成多方面的影响[6 ] . ...
Nutrient limitation and soil development: experimental test of a biogeoche-mical theory.
1
1997
... 近几十年, 人类活动使陆地生态系统的固氮量成倍增加[1 -2 ] , 目前热带、亚热带地区的氮肥使用量占全球使用量的 40%[3 ] , 据预测, 到 2020 年, 全球约2/3的氮输入将发生在热带、亚热带地区[1 ] .我国亚热带地区也存在高氮沉降量现象, 如太湖的常熟生态站大气氮湿沉降量在 2001—2003 年间约为 27 kg/(hm2 ·a)[4 ] , 广东鼎湖山自然保护区在 20 世纪90年代末的降水氮沉降为 38.4 kg/(hm2 ·a)[5 ] .氮沉降的不断增加对我国森林生态系统的结构和功能造成多方面的影响[6 ] . ...
Nitrogenous fertilizers: global distri-bution of consumption and associated emissions of nitrous oxide and ammonia.
1
1994
... 近几十年, 人类活动使陆地生态系统的固氮量成倍增加[1 -2 ] , 目前热带、亚热带地区的氮肥使用量占全球使用量的 40%[3 ] , 据预测, 到 2020 年, 全球约2/3的氮输入将发生在热带、亚热带地区[1 ] .我国亚热带地区也存在高氮沉降量现象, 如太湖的常熟生态站大气氮湿沉降量在 2001—2003 年间约为 27 kg/(hm2 ·a)[4 ] , 广东鼎湖山自然保护区在 20 世纪90年代末的降水氮沉降为 38.4 kg/(hm2 ·a)[5 ] .氮沉降的不断增加对我国森林生态系统的结构和功能造成多方面的影响[6 ] . ...
太湖地区氮素湿沉降动态及生态学意义: 以常熟生态站为例
1
2004
... 近几十年, 人类活动使陆地生态系统的固氮量成倍增加[1 -2 ] , 目前热带、亚热带地区的氮肥使用量占全球使用量的 40%[3 ] , 据预测, 到 2020 年, 全球约2/3的氮输入将发生在热带、亚热带地区[1 ] .我国亚热带地区也存在高氮沉降量现象, 如太湖的常熟生态站大气氮湿沉降量在 2001—2003 年间约为 27 kg/(hm2 ·a)[4 ] , 广东鼎湖山自然保护区在 20 世纪90年代末的降水氮沉降为 38.4 kg/(hm2 ·a)[5 ] .氮沉降的不断增加对我国森林生态系统的结构和功能造成多方面的影响[6 ] . ...
鼎湖山区域大气降水特征和物质元素输入对森林生态系统存在和发育的影响
1
2001
... 近几十年, 人类活动使陆地生态系统的固氮量成倍增加[1 -2 ] , 目前热带、亚热带地区的氮肥使用量占全球使用量的 40%[3 ] , 据预测, 到 2020 年, 全球约2/3的氮输入将发生在热带、亚热带地区[1 ] .我国亚热带地区也存在高氮沉降量现象, 如太湖的常熟生态站大气氮湿沉降量在 2001—2003 年间约为 27 kg/(hm2 ·a)[4 ] , 广东鼎湖山自然保护区在 20 世纪90年代末的降水氮沉降为 38.4 kg/(hm2 ·a)[5 ] .氮沉降的不断增加对我国森林生态系统的结构和功能造成多方面的影响[6 ] . ...
氮沉降对森林植物的影响
2
2003
... 近几十年, 人类活动使陆地生态系统的固氮量成倍增加[1 -2 ] , 目前热带、亚热带地区的氮肥使用量占全球使用量的 40%[3 ] , 据预测, 到 2020 年, 全球约2/3的氮输入将发生在热带、亚热带地区[1 ] .我国亚热带地区也存在高氮沉降量现象, 如太湖的常熟生态站大气氮湿沉降量在 2001—2003 年间约为 27 kg/(hm2 ·a)[4 ] , 广东鼎湖山自然保护区在 20 世纪90年代末的降水氮沉降为 38.4 kg/(hm2 ·a)[5 ] .氮沉降的不断增加对我国森林生态系统的结构和功能造成多方面的影响[6 ] . ...
... 氮添加在总体水平上增加了米槠的全年胸径相对生长速率, 说明研究区内氮素是米槠生长的限制因素.研究区内米槠在不同氮添加水平下的响应不同, 除高氮处理外, 低氮和中氮处理均显著提高了米槠的全年径向生长速率.这与我国目前在热带及亚热带森林生态系统的多数氮沉降试验结果一致, 氮沉降会促进受氮限制的年轻森林中树木的生 长[22 ,31 -33 ] , 树木对氮沉降的响应也受树木种类、研究地点和施肥年限等因素的影响.樊后保等[31 ] 对亚热带杉木的生长研究结果表明, 经过 3 年的施氮处理后, 中氮处理(120 kg N/(hm2 ·a))和高氮处理(240 kg N/(hm2 ·a))均明显促进杉木胸径的生长.在鼎湖山的研究也发现, 中氮处理(100 kg N/(hm2 ·a))使马尾松林胸径生长增加 77.8%[32 ] , 高氮处理则将逐渐对乔木幼苗生长产生负面效应[22 ] .通过对米槠的季节性生长分析发现, 氮添加的生长促进作用主要在水热条件较好的湿润季节产生, 这可能是由于研究地区夏季高温多雨, 土壤氮的周转速率增强, 提高了植物生长过程中氮素的可利用性[34 ] , 当氮素为限制因子时, 额外的氮输入可通过增加土壤和凋落物有效氮[35 ] 、改变基质的 C/N 比[6 ] 及改变土壤微生物组成[36 ] 等方式, 短期提高土壤的氮矿化速率, 进一步提高土壤氮素的可利用性. ...
The globalization of nitrogen deposition: consequences for terrestrial ecosystems.
1
2002
... 乔木生长对于维持森林生态系统功能十分重要, 以往研究表明, 氮沉降的加剧对森林生长和生态系统功能存在不同影响.一方面, 由于陆地生态系统多存在氮限制, 短期内有效氮的增加会促进森林生长, 加速生物量的积累, 从而提高陆地生态系统的固碳量[7 -8 ] , 同时氮添加对树木生长的促进程度还受树龄及不同树种氮利用方式等因素的影 响[9 -10 ] ; 另一方面, 长期施肥实验表明, 过量氮的输入会引起生态系统氮饱和, 导致一系列环境负效应, 如养分失衡、土壤酸化、盐基阳离子流失、有害离子富集和植物抗性下降等.超过生态系统氮负荷之后, 氮添加会抑制植物的生长[11 -12 ] , 引起生态系统衰退[13 -14 ] .国内外研究发现, 温带森林多为氮限制生态系统, 氮沉降可在一定程度上促进森林树木的生长[15 -17 ] .与此同时, 有关氮沉降对热带森林的施肥效果仍存在一定争议, 相对于氮元素, 通常认为热带地区磷对生长的限制作用更强, 一些研究也证实氮沉降对森林生长没有影响[18 -20 ] .然而LeBauer等[21 ] 对 16 个热带森林研究后发现, 部分热带森林类型表现出明显的氮限制特点.由于森林生长对氮沉降的响应存在诸多不确定性[22 -24 ] , 因此对武夷山地区天然米槠林进行氮添加背景下的生长研究有利于评估氮沉降对该区域亚热带常绿阔叶林生态系统的影响. ...
Carbon and nitrogen in Danish forest soils — contents and distri-bution determined by soil order.
1
2003
... 乔木生长对于维持森林生态系统功能十分重要, 以往研究表明, 氮沉降的加剧对森林生长和生态系统功能存在不同影响.一方面, 由于陆地生态系统多存在氮限制, 短期内有效氮的增加会促进森林生长, 加速生物量的积累, 从而提高陆地生态系统的固碳量[7 -8 ] , 同时氮添加对树木生长的促进程度还受树龄及不同树种氮利用方式等因素的影 响[9 -10 ] ; 另一方面, 长期施肥实验表明, 过量氮的输入会引起生态系统氮饱和, 导致一系列环境负效应, 如养分失衡、土壤酸化、盐基阳离子流失、有害离子富集和植物抗性下降等.超过生态系统氮负荷之后, 氮添加会抑制植物的生长[11 -12 ] , 引起生态系统衰退[13 -14 ] .国内外研究发现, 温带森林多为氮限制生态系统, 氮沉降可在一定程度上促进森林树木的生长[15 -17 ] .与此同时, 有关氮沉降对热带森林的施肥效果仍存在一定争议, 相对于氮元素, 通常认为热带地区磷对生长的限制作用更强, 一些研究也证实氮沉降对森林生长没有影响[18 -20 ] .然而LeBauer等[21 ] 对 16 个热带森林研究后发现, 部分热带森林类型表现出明显的氮限制特点.由于森林生长对氮沉降的响应存在诸多不确定性[22 -24 ] , 因此对武夷山地区天然米槠林进行氮添加背景下的生长研究有利于评估氮沉降对该区域亚热带常绿阔叶林生态系统的影响. ...
落叶松人工林的施肥效应
1
2004
... 乔木生长对于维持森林生态系统功能十分重要, 以往研究表明, 氮沉降的加剧对森林生长和生态系统功能存在不同影响.一方面, 由于陆地生态系统多存在氮限制, 短期内有效氮的增加会促进森林生长, 加速生物量的积累, 从而提高陆地生态系统的固碳量[7 -8 ] , 同时氮添加对树木生长的促进程度还受树龄及不同树种氮利用方式等因素的影 响[9 -10 ] ; 另一方面, 长期施肥实验表明, 过量氮的输入会引起生态系统氮饱和, 导致一系列环境负效应, 如养分失衡、土壤酸化、盐基阳离子流失、有害离子富集和植物抗性下降等.超过生态系统氮负荷之后, 氮添加会抑制植物的生长[11 -12 ] , 引起生态系统衰退[13 -14 ] .国内外研究发现, 温带森林多为氮限制生态系统, 氮沉降可在一定程度上促进森林树木的生长[15 -17 ] .与此同时, 有关氮沉降对热带森林的施肥效果仍存在一定争议, 相对于氮元素, 通常认为热带地区磷对生长的限制作用更强, 一些研究也证实氮沉降对森林生长没有影响[18 -20 ] .然而LeBauer等[21 ] 对 16 个热带森林研究后发现, 部分热带森林类型表现出明显的氮限制特点.由于森林生长对氮沉降的响应存在诸多不确定性[22 -24 ] , 因此对武夷山地区天然米槠林进行氮添加背景下的生长研究有利于评估氮沉降对该区域亚热带常绿阔叶林生态系统的影响. ...
Impact of long-term nitrogen addition on carbon stocks in trees and soils in northern Europe.
1
2008
... 乔木生长对于维持森林生态系统功能十分重要, 以往研究表明, 氮沉降的加剧对森林生长和生态系统功能存在不同影响.一方面, 由于陆地生态系统多存在氮限制, 短期内有效氮的增加会促进森林生长, 加速生物量的积累, 从而提高陆地生态系统的固碳量[7 -8 ] , 同时氮添加对树木生长的促进程度还受树龄及不同树种氮利用方式等因素的影 响[9 -10 ] ; 另一方面, 长期施肥实验表明, 过量氮的输入会引起生态系统氮饱和, 导致一系列环境负效应, 如养分失衡、土壤酸化、盐基阳离子流失、有害离子富集和植物抗性下降等.超过生态系统氮负荷之后, 氮添加会抑制植物的生长[11 -12 ] , 引起生态系统衰退[13 -14 ] .国内外研究发现, 温带森林多为氮限制生态系统, 氮沉降可在一定程度上促进森林树木的生长[15 -17 ] .与此同时, 有关氮沉降对热带森林的施肥效果仍存在一定争议, 相对于氮元素, 通常认为热带地区磷对生长的限制作用更强, 一些研究也证实氮沉降对森林生长没有影响[18 -20 ] .然而LeBauer等[21 ] 对 16 个热带森林研究后发现, 部分热带森林类型表现出明显的氮限制特点.由于森林生长对氮沉降的响应存在诸多不确定性[22 -24 ] , 因此对武夷山地区天然米槠林进行氮添加背景下的生长研究有利于评估氮沉降对该区域亚热带常绿阔叶林生态系统的影响. ...
Increased tree carbon storage in response to nitrogen deposition in the US.
1
2010
... 乔木生长对于维持森林生态系统功能十分重要, 以往研究表明, 氮沉降的加剧对森林生长和生态系统功能存在不同影响.一方面, 由于陆地生态系统多存在氮限制, 短期内有效氮的增加会促进森林生长, 加速生物量的积累, 从而提高陆地生态系统的固碳量[7 -8 ] , 同时氮添加对树木生长的促进程度还受树龄及不同树种氮利用方式等因素的影 响[9 -10 ] ; 另一方面, 长期施肥实验表明, 过量氮的输入会引起生态系统氮饱和, 导致一系列环境负效应, 如养分失衡、土壤酸化、盐基阳离子流失、有害离子富集和植物抗性下降等.超过生态系统氮负荷之后, 氮添加会抑制植物的生长[11 -12 ] , 引起生态系统衰退[13 -14 ] .国内外研究发现, 温带森林多为氮限制生态系统, 氮沉降可在一定程度上促进森林树木的生长[15 -17 ] .与此同时, 有关氮沉降对热带森林的施肥效果仍存在一定争议, 相对于氮元素, 通常认为热带地区磷对生长的限制作用更强, 一些研究也证实氮沉降对森林生长没有影响[18 -20 ] .然而LeBauer等[21 ] 对 16 个热带森林研究后发现, 部分热带森林类型表现出明显的氮限制特点.由于森林生长对氮沉降的响应存在诸多不确定性[22 -24 ] , 因此对武夷山地区天然米槠林进行氮添加背景下的生长研究有利于评估氮沉降对该区域亚热带常绿阔叶林生态系统的影响. ...
McDowell W, Nadelhoffer K, et al. Nitrogen saturation in temperate forest ecosystems-Hypotheses revisited.
1
1998
... 乔木生长对于维持森林生态系统功能十分重要, 以往研究表明, 氮沉降的加剧对森林生长和生态系统功能存在不同影响.一方面, 由于陆地生态系统多存在氮限制, 短期内有效氮的增加会促进森林生长, 加速生物量的积累, 从而提高陆地生态系统的固碳量[7 -8 ] , 同时氮添加对树木生长的促进程度还受树龄及不同树种氮利用方式等因素的影 响[9 -10 ] ; 另一方面, 长期施肥实验表明, 过量氮的输入会引起生态系统氮饱和, 导致一系列环境负效应, 如养分失衡、土壤酸化、盐基阳离子流失、有害离子富集和植物抗性下降等.超过生态系统氮负荷之后, 氮添加会抑制植物的生长[11 -12 ] , 引起生态系统衰退[13 -14 ] .国内外研究发现, 温带森林多为氮限制生态系统, 氮沉降可在一定程度上促进森林树木的生长[15 -17 ] .与此同时, 有关氮沉降对热带森林的施肥效果仍存在一定争议, 相对于氮元素, 通常认为热带地区磷对生长的限制作用更强, 一些研究也证实氮沉降对森林生长没有影响[18 -20 ] .然而LeBauer等[21 ] 对 16 个热带森林研究后发现, 部分热带森林类型表现出明显的氮限制特点.由于森林生长对氮沉降的响应存在诸多不确定性[22 -24 ] , 因此对武夷山地区天然米槠林进行氮添加背景下的生长研究有利于评估氮沉降对该区域亚热带常绿阔叶林生态系统的影响. ...
Variable effects of nitrogen additions on the stability and turnover of soil C.
1
2002
... 乔木生长对于维持森林生态系统功能十分重要, 以往研究表明, 氮沉降的加剧对森林生长和生态系统功能存在不同影响.一方面, 由于陆地生态系统多存在氮限制, 短期内有效氮的增加会促进森林生长, 加速生物量的积累, 从而提高陆地生态系统的固碳量[7 -8 ] , 同时氮添加对树木生长的促进程度还受树龄及不同树种氮利用方式等因素的影 响[9 -10 ] ; 另一方面, 长期施肥实验表明, 过量氮的输入会引起生态系统氮饱和, 导致一系列环境负效应, 如养分失衡、土壤酸化、盐基阳离子流失、有害离子富集和植物抗性下降等.超过生态系统氮负荷之后, 氮添加会抑制植物的生长[11 -12 ] , 引起生态系统衰退[13 -14 ] .国内外研究发现, 温带森林多为氮限制生态系统, 氮沉降可在一定程度上促进森林树木的生长[15 -17 ] .与此同时, 有关氮沉降对热带森林的施肥效果仍存在一定争议, 相对于氮元素, 通常认为热带地区磷对生长的限制作用更强, 一些研究也证实氮沉降对森林生长没有影响[18 -20 ] .然而LeBauer等[21 ] 对 16 个热带森林研究后发现, 部分热带森林类型表现出明显的氮限制特点.由于森林生长对氮沉降的响应存在诸多不确定性[22 -24 ] , 因此对武夷山地区天然米槠林进行氮添加背景下的生长研究有利于评估氮沉降对该区域亚热带常绿阔叶林生态系统的影响. ...
Nitrogen deposition and the biodiversity of boreal forests: implications for the nitrogen critical load.
1
2005
... 乔木生长对于维持森林生态系统功能十分重要, 以往研究表明, 氮沉降的加剧对森林生长和生态系统功能存在不同影响.一方面, 由于陆地生态系统多存在氮限制, 短期内有效氮的增加会促进森林生长, 加速生物量的积累, 从而提高陆地生态系统的固碳量[7 -8 ] , 同时氮添加对树木生长的促进程度还受树龄及不同树种氮利用方式等因素的影 响[9 -10 ] ; 另一方面, 长期施肥实验表明, 过量氮的输入会引起生态系统氮饱和, 导致一系列环境负效应, 如养分失衡、土壤酸化、盐基阳离子流失、有害离子富集和植物抗性下降等.超过生态系统氮负荷之后, 氮添加会抑制植物的生长[11 -12 ] , 引起生态系统衰退[13 -14 ] .国内外研究发现, 温带森林多为氮限制生态系统, 氮沉降可在一定程度上促进森林树木的生长[15 -17 ] .与此同时, 有关氮沉降对热带森林的施肥效果仍存在一定争议, 相对于氮元素, 通常认为热带地区磷对生长的限制作用更强, 一些研究也证实氮沉降对森林生长没有影响[18 -20 ] .然而LeBauer等[21 ] 对 16 个热带森林研究后发现, 部分热带森林类型表现出明显的氮限制特点.由于森林生长对氮沉降的响应存在诸多不确定性[22 -24 ] , 因此对武夷山地区天然米槠林进行氮添加背景下的生长研究有利于评估氮沉降对该区域亚热带常绿阔叶林生态系统的影响. ...
Analyses of the impact of changes in atmospheric deposition and climate on forest growth in European monitoring plots: a stand growth approach.
1
2009
... 乔木生长对于维持森林生态系统功能十分重要, 以往研究表明, 氮沉降的加剧对森林生长和生态系统功能存在不同影响.一方面, 由于陆地生态系统多存在氮限制, 短期内有效氮的增加会促进森林生长, 加速生物量的积累, 从而提高陆地生态系统的固碳量[7 -8 ] , 同时氮添加对树木生长的促进程度还受树龄及不同树种氮利用方式等因素的影 响[9 -10 ] ; 另一方面, 长期施肥实验表明, 过量氮的输入会引起生态系统氮饱和, 导致一系列环境负效应, 如养分失衡、土壤酸化、盐基阳离子流失、有害离子富集和植物抗性下降等.超过生态系统氮负荷之后, 氮添加会抑制植物的生长[11 -12 ] , 引起生态系统衰退[13 -14 ] .国内外研究发现, 温带森林多为氮限制生态系统, 氮沉降可在一定程度上促进森林树木的生长[15 -17 ] .与此同时, 有关氮沉降对热带森林的施肥效果仍存在一定争议, 相对于氮元素, 通常认为热带地区磷对生长的限制作用更强, 一些研究也证实氮沉降对森林生长没有影响[18 -20 ] .然而LeBauer等[21 ] 对 16 个热带森林研究后发现, 部分热带森林类型表现出明显的氮限制特点.由于森林生长对氮沉降的响应存在诸多不确定性[22 -24 ] , 因此对武夷山地区天然米槠林进行氮添加背景下的生长研究有利于评估氮沉降对该区域亚热带常绿阔叶林生态系统的影响. ...
Short and long-term impacts of nitrogen deposition on carbon sequestration by forest ecosystems
0
2014
The impact of atmospheric deposition and climate on forest growth in European monitoring plots: an individual tree growth model.
1
2009
... 乔木生长对于维持森林生态系统功能十分重要, 以往研究表明, 氮沉降的加剧对森林生长和生态系统功能存在不同影响.一方面, 由于陆地生态系统多存在氮限制, 短期内有效氮的增加会促进森林生长, 加速生物量的积累, 从而提高陆地生态系统的固碳量[7 -8 ] , 同时氮添加对树木生长的促进程度还受树龄及不同树种氮利用方式等因素的影 响[9 -10 ] ; 另一方面, 长期施肥实验表明, 过量氮的输入会引起生态系统氮饱和, 导致一系列环境负效应, 如养分失衡、土壤酸化、盐基阳离子流失、有害离子富集和植物抗性下降等.超过生态系统氮负荷之后, 氮添加会抑制植物的生长[11 -12 ] , 引起生态系统衰退[13 -14 ] .国内外研究发现, 温带森林多为氮限制生态系统, 氮沉降可在一定程度上促进森林树木的生长[15 -17 ] .与此同时, 有关氮沉降对热带森林的施肥效果仍存在一定争议, 相对于氮元素, 通常认为热带地区磷对生长的限制作用更强, 一些研究也证实氮沉降对森林生长没有影响[18 -20 ] .然而LeBauer等[21 ] 对 16 个热带森林研究后发现, 部分热带森林类型表现出明显的氮限制特点.由于森林生长对氮沉降的响应存在诸多不确定性[22 -24 ] , 因此对武夷山地区天然米槠林进行氮添加背景下的生长研究有利于评估氮沉降对该区域亚热带常绿阔叶林生态系统的影响. ...
Effects of nitrogen additions on above- and belowground carbon dynamics in two tropical forests.
1
2011
... 乔木生长对于维持森林生态系统功能十分重要, 以往研究表明, 氮沉降的加剧对森林生长和生态系统功能存在不同影响.一方面, 由于陆地生态系统多存在氮限制, 短期内有效氮的增加会促进森林生长, 加速生物量的积累, 从而提高陆地生态系统的固碳量[7 -8 ] , 同时氮添加对树木生长的促进程度还受树龄及不同树种氮利用方式等因素的影 响[9 -10 ] ; 另一方面, 长期施肥实验表明, 过量氮的输入会引起生态系统氮饱和, 导致一系列环境负效应, 如养分失衡、土壤酸化、盐基阳离子流失、有害离子富集和植物抗性下降等.超过生态系统氮负荷之后, 氮添加会抑制植物的生长[11 -12 ] , 引起生态系统衰退[13 -14 ] .国内外研究发现, 温带森林多为氮限制生态系统, 氮沉降可在一定程度上促进森林树木的生长[15 -17 ] .与此同时, 有关氮沉降对热带森林的施肥效果仍存在一定争议, 相对于氮元素, 通常认为热带地区磷对生长的限制作用更强, 一些研究也证实氮沉降对森林生长没有影响[18 -20 ] .然而LeBauer等[21 ] 对 16 个热带森林研究后发现, 部分热带森林类型表现出明显的氮限制特点.由于森林生长对氮沉降的响应存在诸多不确定性[22 -24 ] , 因此对武夷山地区天然米槠林进行氮添加背景下的生长研究有利于评估氮沉降对该区域亚热带常绿阔叶林生态系统的影响. ...
Effects of nitrogen andphosphorus fertilization in a lowland evergreen rainforest.
0
1999
Terre-strial phosphorus limitation: mechanisms, implications, and nitrogen-phosphorus interactions.
1
2010
... 乔木生长对于维持森林生态系统功能十分重要, 以往研究表明, 氮沉降的加剧对森林生长和生态系统功能存在不同影响.一方面, 由于陆地生态系统多存在氮限制, 短期内有效氮的增加会促进森林生长, 加速生物量的积累, 从而提高陆地生态系统的固碳量[7 -8 ] , 同时氮添加对树木生长的促进程度还受树龄及不同树种氮利用方式等因素的影 响[9 -10 ] ; 另一方面, 长期施肥实验表明, 过量氮的输入会引起生态系统氮饱和, 导致一系列环境负效应, 如养分失衡、土壤酸化、盐基阳离子流失、有害离子富集和植物抗性下降等.超过生态系统氮负荷之后, 氮添加会抑制植物的生长[11 -12 ] , 引起生态系统衰退[13 -14 ] .国内外研究发现, 温带森林多为氮限制生态系统, 氮沉降可在一定程度上促进森林树木的生长[15 -17 ] .与此同时, 有关氮沉降对热带森林的施肥效果仍存在一定争议, 相对于氮元素, 通常认为热带地区磷对生长的限制作用更强, 一些研究也证实氮沉降对森林生长没有影响[18 -20 ] .然而LeBauer等[21 ] 对 16 个热带森林研究后发现, 部分热带森林类型表现出明显的氮限制特点.由于森林生长对氮沉降的响应存在诸多不确定性[22 -24 ] , 因此对武夷山地区天然米槠林进行氮添加背景下的生长研究有利于评估氮沉降对该区域亚热带常绿阔叶林生态系统的影响. ...
Nitrogen limitation of net primary productivity in terrestrial ecosystems is globally distributed.
1
2008
... 乔木生长对于维持森林生态系统功能十分重要, 以往研究表明, 氮沉降的加剧对森林生长和生态系统功能存在不同影响.一方面, 由于陆地生态系统多存在氮限制, 短期内有效氮的增加会促进森林生长, 加速生物量的积累, 从而提高陆地生态系统的固碳量[7 -8 ] , 同时氮添加对树木生长的促进程度还受树龄及不同树种氮利用方式等因素的影 响[9 -10 ] ; 另一方面, 长期施肥实验表明, 过量氮的输入会引起生态系统氮饱和, 导致一系列环境负效应, 如养分失衡、土壤酸化、盐基阳离子流失、有害离子富集和植物抗性下降等.超过生态系统氮负荷之后, 氮添加会抑制植物的生长[11 -12 ] , 引起生态系统衰退[13 -14 ] .国内外研究发现, 温带森林多为氮限制生态系统, 氮沉降可在一定程度上促进森林树木的生长[15 -17 ] .与此同时, 有关氮沉降对热带森林的施肥效果仍存在一定争议, 相对于氮元素, 通常认为热带地区磷对生长的限制作用更强, 一些研究也证实氮沉降对森林生长没有影响[18 -20 ] .然而LeBauer等[21 ] 对 16 个热带森林研究后发现, 部分热带森林类型表现出明显的氮限制特点.由于森林生长对氮沉降的响应存在诸多不确定性[22 -24 ] , 因此对武夷山地区天然米槠林进行氮添加背景下的生长研究有利于评估氮沉降对该区域亚热带常绿阔叶林生态系统的影响. ...
模拟氮沉降对南亚热带两种乔木幼苗生物量及其分配的影响
3
2005
... 乔木生长对于维持森林生态系统功能十分重要, 以往研究表明, 氮沉降的加剧对森林生长和生态系统功能存在不同影响.一方面, 由于陆地生态系统多存在氮限制, 短期内有效氮的增加会促进森林生长, 加速生物量的积累, 从而提高陆地生态系统的固碳量[7 -8 ] , 同时氮添加对树木生长的促进程度还受树龄及不同树种氮利用方式等因素的影 响[9 -10 ] ; 另一方面, 长期施肥实验表明, 过量氮的输入会引起生态系统氮饱和, 导致一系列环境负效应, 如养分失衡、土壤酸化、盐基阳离子流失、有害离子富集和植物抗性下降等.超过生态系统氮负荷之后, 氮添加会抑制植物的生长[11 -12 ] , 引起生态系统衰退[13 -14 ] .国内外研究发现, 温带森林多为氮限制生态系统, 氮沉降可在一定程度上促进森林树木的生长[15 -17 ] .与此同时, 有关氮沉降对热带森林的施肥效果仍存在一定争议, 相对于氮元素, 通常认为热带地区磷对生长的限制作用更强, 一些研究也证实氮沉降对森林生长没有影响[18 -20 ] .然而LeBauer等[21 ] 对 16 个热带森林研究后发现, 部分热带森林类型表现出明显的氮限制特点.由于森林生长对氮沉降的响应存在诸多不确定性[22 -24 ] , 因此对武夷山地区天然米槠林进行氮添加背景下的生长研究有利于评估氮沉降对该区域亚热带常绿阔叶林生态系统的影响. ...
... 氮添加在总体水平上增加了米槠的全年胸径相对生长速率, 说明研究区内氮素是米槠生长的限制因素.研究区内米槠在不同氮添加水平下的响应不同, 除高氮处理外, 低氮和中氮处理均显著提高了米槠的全年径向生长速率.这与我国目前在热带及亚热带森林生态系统的多数氮沉降试验结果一致, 氮沉降会促进受氮限制的年轻森林中树木的生 长[22 ,31 -33 ] , 树木对氮沉降的响应也受树木种类、研究地点和施肥年限等因素的影响.樊后保等[31 ] 对亚热带杉木的生长研究结果表明, 经过 3 年的施氮处理后, 中氮处理(120 kg N/(hm2 ·a))和高氮处理(240 kg N/(hm2 ·a))均明显促进杉木胸径的生长.在鼎湖山的研究也发现, 中氮处理(100 kg N/(hm2 ·a))使马尾松林胸径生长增加 77.8%[32 ] , 高氮处理则将逐渐对乔木幼苗生长产生负面效应[22 ] .通过对米槠的季节性生长分析发现, 氮添加的生长促进作用主要在水热条件较好的湿润季节产生, 这可能是由于研究地区夏季高温多雨, 土壤氮的周转速率增强, 提高了植物生长过程中氮素的可利用性[34 ] , 当氮素为限制因子时, 额外的氮输入可通过增加土壤和凋落物有效氮[35 ] 、改变基质的 C/N 比[6 ] 及改变土壤微生物组成[36 ] 等方式, 短期提高土壤的氮矿化速率, 进一步提高土壤氮素的可利用性. ...
... [22 ].通过对米槠的季节性生长分析发现, 氮添加的生长促进作用主要在水热条件较好的湿润季节产生, 这可能是由于研究地区夏季高温多雨, 土壤氮的周转速率增强, 提高了植物生长过程中氮素的可利用性[34 ] , 当氮素为限制因子时, 额外的氮输入可通过增加土壤和凋落物有效氮[35 ] 、改变基质的 C/N 比[6 ] 及改变土壤微生物组成[36 ] 等方式, 短期提高土壤的氮矿化速率, 进一步提高土壤氮素的可利用性. ...
模拟氮沉降对三种南亚热带树苗生长和光合作用的影响
0
2004
模拟降水变化和土壤施氮对浙江古田山 5 个树种幼苗生长和生物量的影响
1
2011
... 乔木生长对于维持森林生态系统功能十分重要, 以往研究表明, 氮沉降的加剧对森林生长和生态系统功能存在不同影响.一方面, 由于陆地生态系统多存在氮限制, 短期内有效氮的增加会促进森林生长, 加速生物量的积累, 从而提高陆地生态系统的固碳量[7 -8 ] , 同时氮添加对树木生长的促进程度还受树龄及不同树种氮利用方式等因素的影 响[9 -10 ] ; 另一方面, 长期施肥实验表明, 过量氮的输入会引起生态系统氮饱和, 导致一系列环境负效应, 如养分失衡、土壤酸化、盐基阳离子流失、有害离子富集和植物抗性下降等.超过生态系统氮负荷之后, 氮添加会抑制植物的生长[11 -12 ] , 引起生态系统衰退[13 -14 ] .国内外研究发现, 温带森林多为氮限制生态系统, 氮沉降可在一定程度上促进森林树木的生长[15 -17 ] .与此同时, 有关氮沉降对热带森林的施肥效果仍存在一定争议, 相对于氮元素, 通常认为热带地区磷对生长的限制作用更强, 一些研究也证实氮沉降对森林生长没有影响[18 -20 ] .然而LeBauer等[21 ] 对 16 个热带森林研究后发现, 部分热带森林类型表现出明显的氮限制特点.由于森林生长对氮沉降的响应存在诸多不确定性[22 -24 ] , 因此对武夷山地区天然米槠林进行氮添加背景下的生长研究有利于评估氮沉降对该区域亚热带常绿阔叶林生态系统的影响. ...
武夷山国家级自然保护区土壤的现状与保护
2
... 研究区位于福建武夷山国家自然保护区, 地处武夷山脉北端, 武夷山市、建阳市、光泽县和邵武市四县(市)交界处, 地理范围为 117°27′—51′E, 27°33′—54′N.该区属典型的亚热带季风气候, 夏季高温多雨, 冬季温和湿润.年均气温 8.5~18°C, 年均相对湿度 78%~84%, 年均降水量 1486~2150 mm, 无霜期 253~273 天, 年平均雾日达 120 天[25 ] .研究区内地带性植被为亚热常绿阔叶林, 植被类型属于保存比较完好的米槠群落, 物种丰富, 层次分明[26 ] , 地带性土壤为红黄壤[25 ] . ...
... [25 ]. ...
武夷山国家级自然保护区森林土壤涵水量估测及空间分析
1
2014
... 研究区位于福建武夷山国家自然保护区, 地处武夷山脉北端, 武夷山市、建阳市、光泽县和邵武市四县(市)交界处, 地理范围为 117°27′—51′E, 27°33′—54′N.该区属典型的亚热带季风气候, 夏季高温多雨, 冬季温和湿润.年均气温 8.5~18°C, 年均相对湿度 78%~84%, 年均降水量 1486~2150 mm, 无霜期 253~273 天, 年平均雾日达 120 天[25 ] .研究区内地带性植被为亚热常绿阔叶林, 植被类型属于保存比较完好的米槠群落, 物种丰富, 层次分明[26 ] , 地带性土壤为红黄壤[25 ] . ...
The use of precision dendrometers in research on daily stem size and wood property variation: a review.
1
2009
... 对样方内所有胸径(diameter at breath height, DBH) ≥5 cm的米槠于1.3 m高处安装自制生长环, 并测定其初始胸径值.2014年1月至12月, 每月的月末用电子游标卡尺测量其胸围变化量.同时, 在每种处理水平下选取1棵干形较直、胸径及树高均接近样地内同物种平均值的米槠作为标准树(表2 ), 安装自动胸径记录仪(Dendrometer-DRL26, 捷克), 每隔 30 分钟记录 1 次数据, 对采集的数据用USB 红外转输下载.树木径向生长测量仪是一种可以自动记录树木原位生长精确数据的电子仪器, 与传统的监测方法(直接测量胸径)相比, 该仪器能够识别树干水分引起的日际收缩和膨胀波动[27 ] .本研究共安置 4 个带状记录仪, 自动记录生长数据.采用的树木生长数据来源于自制生长环法和自动记录仪法两部分, 时间范围为 2014 年 1—12月. ...
Dendrometer and intra-annual tree growth: what kind of information can be inferred.
3
2007
... 依据 Deslauriers 等[28 ] 的方法, 结合实验样地的经纬度及海拔, 树木的生长时期由连续 15 日的径向日变化均值确定, 该方法比直接观察胸径值变化趋势更准确.2014 年 3 月 15 日至 2015 年 3 月 1日, 使用单样本t 检验对每连续15日的径向日变化均值(即连续15日的每日径向变化值取平均)与0做比较, 当值的分布显著大于零时, 认为开始发生明显的径向生长[29 ] .树木的径向日变化量由Bouriaud等[30 ] 提出的最大值法计算而来, 从记录米槠茎干每日生长的 48 个原始数据中提取日最大值, 计算连续两日的最大值之差, 求得树木茎干的日变化量. ...
... 图 1显示米槠全年的茎干累计生长变化动态, 可以看到4棵样树的全年径向生长表现出一致的变化规律, 冬季及初春茎干增长极为缓慢, 夏季表现为快速的持续增长, 随后生长速率立刻放缓.使用Deslauriers 等[28 ] 判断树木不同生长阶段的方法, 对每连续 15 天的日径向变化值进行分析, 结果如图2 所示.对4棵样树分别进行分析, 发现第一个与 0 有显著差异的正值均出现在4月中旬至5 月中旬.此阶段为茎干水分恢复阶段(5 月), 树木茎干表现为因吸水而出现的可逆性逐渐增粗.之后, 样树 a和 d 在 6 月初出现一个与 0 无显著差异的值, 此阶段为水分恢复与茎干生长之间的平台期.在 10—11月, 4棵样树先后出现最后一个与0有显著差异的正值.因此, 可将 6—10 月划分为茎干快速生长阶段, 树木茎干在此阶段快速增粗, 径向增长加快.11—4 月则为茎干缓慢生长阶段, 此阶段没有出现与0有显著差异的正值, 米槠茎干径向生长速率大幅放缓. ...
... 选择合适的时间尺度对自动记录数据进行分析, 可以更准确地识别树木的生长阶段.Deslau-riers 等[28 ] 认为, 在较低海拔地区, 选择 10 日以上的时间尺度, 较易识别树木茎干生长的主要阶段.根据样地所处的位置以及海拔, 本研究选择 15 日的时间尺度以及 4 株样树来识别米槠全年的不同生长阶段.结果表明, 样树a和d的径向生长在6月初至6月中旬有一段时间表现为非明显增长的平台期(图 2 ), 以此来区分树木在本年内的快速生长阶段.样树b和c未识别出这一时期, 可能主要是因为亚热带地区春季降雨量大且持续时间较长, 树木的补水阶段与夏季的径向快速生长阶段实现快速过渡. ...
小兴安岭红松日径向变化及其对气象因子的响应
1
2014
... 依据 Deslauriers 等[28 ] 的方法, 结合实验样地的经纬度及海拔, 树木的生长时期由连续 15 日的径向日变化均值确定, 该方法比直接观察胸径值变化趋势更准确.2014 年 3 月 15 日至 2015 年 3 月 1日, 使用单样本t 检验对每连续15日的径向日变化均值(即连续15日的每日径向变化值取平均)与0做比较, 当值的分布显著大于零时, 认为开始发生明显的径向生长[29 ] .树木的径向日变化量由Bouriaud等[30 ] 提出的最大值法计算而来, 从记录米槠茎干每日生长的 48 个原始数据中提取日最大值, 计算连续两日的最大值之差, 求得树木茎干的日变化量. ...
Intra-annual variations in climate influence growth and wood density of Norway spruce.
1
2005
... 依据 Deslauriers 等[28 ] 的方法, 结合实验样地的经纬度及海拔, 树木的生长时期由连续 15 日的径向日变化均值确定, 该方法比直接观察胸径值变化趋势更准确.2014 年 3 月 15 日至 2015 年 3 月 1日, 使用单样本t 检验对每连续15日的径向日变化均值(即连续15日的每日径向变化值取平均)与0做比较, 当值的分布显著大于零时, 认为开始发生明显的径向生长[29 ] .树木的径向日变化量由Bouriaud等[30 ] 提出的最大值法计算而来, 从记录米槠茎干每日生长的 48 个原始数据中提取日最大值, 计算连续两日的最大值之差, 求得树木茎干的日变化量. ...
亚热带杉木(Cun-ninghamia lanceolata )人工林生长与土壤养分对氮沉降的响应
2
2007
... 氮添加在总体水平上增加了米槠的全年胸径相对生长速率, 说明研究区内氮素是米槠生长的限制因素.研究区内米槠在不同氮添加水平下的响应不同, 除高氮处理外, 低氮和中氮处理均显著提高了米槠的全年径向生长速率.这与我国目前在热带及亚热带森林生态系统的多数氮沉降试验结果一致, 氮沉降会促进受氮限制的年轻森林中树木的生 长[22 ,31 -33 ] , 树木对氮沉降的响应也受树木种类、研究地点和施肥年限等因素的影响.樊后保等[31 ] 对亚热带杉木的生长研究结果表明, 经过 3 年的施氮处理后, 中氮处理(120 kg N/(hm2 ·a))和高氮处理(240 kg N/(hm2 ·a))均明显促进杉木胸径的生长.在鼎湖山的研究也发现, 中氮处理(100 kg N/(hm2 ·a))使马尾松林胸径生长增加 77.8%[32 ] , 高氮处理则将逐渐对乔木幼苗生长产生负面效应[22 ] .通过对米槠的季节性生长分析发现, 氮添加的生长促进作用主要在水热条件较好的湿润季节产生, 这可能是由于研究地区夏季高温多雨, 土壤氮的周转速率增强, 提高了植物生长过程中氮素的可利用性[34 ] , 当氮素为限制因子时, 额外的氮输入可通过增加土壤和凋落物有效氮[35 ] 、改变基质的 C/N 比[6 ] 及改变土壤微生物组成[36 ] 等方式, 短期提高土壤的氮矿化速率, 进一步提高土壤氮素的可利用性. ...
... [31 ]对亚热带杉木的生长研究结果表明, 经过 3 年的施氮处理后, 中氮处理(120 kg N/(hm2 ·a))和高氮处理(240 kg N/(hm2 ·a))均明显促进杉木胸径的生长.在鼎湖山的研究也发现, 中氮处理(100 kg N/(hm2 ·a))使马尾松林胸径生长增加 77.8%[32 ] , 高氮处理则将逐渐对乔木幼苗生长产生负面效应[22 ] .通过对米槠的季节性生长分析发现, 氮添加的生长促进作用主要在水热条件较好的湿润季节产生, 这可能是由于研究地区夏季高温多雨, 土壤氮的周转速率增强, 提高了植物生长过程中氮素的可利用性[34 ] , 当氮素为限制因子时, 额外的氮输入可通过增加土壤和凋落物有效氮[35 ] 、改变基质的 C/N 比[6 ] 及改变土壤微生物组成[36 ] 等方式, 短期提高土壤的氮矿化速率, 进一步提高土壤氮素的可利用性. ...
鼎湖山主要森林类型植物胸径生长对氮沉降增加的初期响应
1
2005
... 氮添加在总体水平上增加了米槠的全年胸径相对生长速率, 说明研究区内氮素是米槠生长的限制因素.研究区内米槠在不同氮添加水平下的响应不同, 除高氮处理外, 低氮和中氮处理均显著提高了米槠的全年径向生长速率.这与我国目前在热带及亚热带森林生态系统的多数氮沉降试验结果一致, 氮沉降会促进受氮限制的年轻森林中树木的生 长[22 ,31 -33 ] , 树木对氮沉降的响应也受树木种类、研究地点和施肥年限等因素的影响.樊后保等[31 ] 对亚热带杉木的生长研究结果表明, 经过 3 年的施氮处理后, 中氮处理(120 kg N/(hm2 ·a))和高氮处理(240 kg N/(hm2 ·a))均明显促进杉木胸径的生长.在鼎湖山的研究也发现, 中氮处理(100 kg N/(hm2 ·a))使马尾松林胸径生长增加 77.8%[32 ] , 高氮处理则将逐渐对乔木幼苗生长产生负面效应[22 ] .通过对米槠的季节性生长分析发现, 氮添加的生长促进作用主要在水热条件较好的湿润季节产生, 这可能是由于研究地区夏季高温多雨, 土壤氮的周转速率增强, 提高了植物生长过程中氮素的可利用性[34 ] , 当氮素为限制因子时, 额外的氮输入可通过增加土壤和凋落物有效氮[35 ] 、改变基质的 C/N 比[6 ] 及改变土壤微生物组成[36 ] 等方式, 短期提高土壤的氮矿化速率, 进一步提高土壤氮素的可利用性. ...
氮磷添加对海南热带山地雨林碳循环的影响[D]
1
2013
... 氮添加在总体水平上增加了米槠的全年胸径相对生长速率, 说明研究区内氮素是米槠生长的限制因素.研究区内米槠在不同氮添加水平下的响应不同, 除高氮处理外, 低氮和中氮处理均显著提高了米槠的全年径向生长速率.这与我国目前在热带及亚热带森林生态系统的多数氮沉降试验结果一致, 氮沉降会促进受氮限制的年轻森林中树木的生 长[22 ,31 -33 ] , 树木对氮沉降的响应也受树木种类、研究地点和施肥年限等因素的影响.樊后保等[31 ] 对亚热带杉木的生长研究结果表明, 经过 3 年的施氮处理后, 中氮处理(120 kg N/(hm2 ·a))和高氮处理(240 kg N/(hm2 ·a))均明显促进杉木胸径的生长.在鼎湖山的研究也发现, 中氮处理(100 kg N/(hm2 ·a))使马尾松林胸径生长增加 77.8%[32 ] , 高氮处理则将逐渐对乔木幼苗生长产生负面效应[22 ] .通过对米槠的季节性生长分析发现, 氮添加的生长促进作用主要在水热条件较好的湿润季节产生, 这可能是由于研究地区夏季高温多雨, 土壤氮的周转速率增强, 提高了植物生长过程中氮素的可利用性[34 ] , 当氮素为限制因子时, 额外的氮输入可通过增加土壤和凋落物有效氮[35 ] 、改变基质的 C/N 比[6 ] 及改变土壤微生物组成[36 ] 等方式, 短期提高土壤的氮矿化速率, 进一步提高土壤氮素的可利用性. ...
Effect of simulated acid rain on potential carbon and nitrogen mineralization in forest soils.
1
2008
... 氮添加在总体水平上增加了米槠的全年胸径相对生长速率, 说明研究区内氮素是米槠生长的限制因素.研究区内米槠在不同氮添加水平下的响应不同, 除高氮处理外, 低氮和中氮处理均显著提高了米槠的全年径向生长速率.这与我国目前在热带及亚热带森林生态系统的多数氮沉降试验结果一致, 氮沉降会促进受氮限制的年轻森林中树木的生 长[22 ,31 -33 ] , 树木对氮沉降的响应也受树木种类、研究地点和施肥年限等因素的影响.樊后保等[31 ] 对亚热带杉木的生长研究结果表明, 经过 3 年的施氮处理后, 中氮处理(120 kg N/(hm2 ·a))和高氮处理(240 kg N/(hm2 ·a))均明显促进杉木胸径的生长.在鼎湖山的研究也发现, 中氮处理(100 kg N/(hm2 ·a))使马尾松林胸径生长增加 77.8%[32 ] , 高氮处理则将逐渐对乔木幼苗生长产生负面效应[22 ] .通过对米槠的季节性生长分析发现, 氮添加的生长促进作用主要在水热条件较好的湿润季节产生, 这可能是由于研究地区夏季高温多雨, 土壤氮的周转速率增强, 提高了植物生长过程中氮素的可利用性[34 ] , 当氮素为限制因子时, 额外的氮输入可通过增加土壤和凋落物有效氮[35 ] 、改变基质的 C/N 比[6 ] 及改变土壤微生物组成[36 ] 等方式, 短期提高土壤的氮矿化速率, 进一步提高土壤氮素的可利用性. ...
Impact of nitrogen deposition on nitrogen cycling in forests: a synthesis of NITREX data.
1
1998
... 氮添加在总体水平上增加了米槠的全年胸径相对生长速率, 说明研究区内氮素是米槠生长的限制因素.研究区内米槠在不同氮添加水平下的响应不同, 除高氮处理外, 低氮和中氮处理均显著提高了米槠的全年径向生长速率.这与我国目前在热带及亚热带森林生态系统的多数氮沉降试验结果一致, 氮沉降会促进受氮限制的年轻森林中树木的生 长[22 ,31 -33 ] , 树木对氮沉降的响应也受树木种类、研究地点和施肥年限等因素的影响.樊后保等[31 ] 对亚热带杉木的生长研究结果表明, 经过 3 年的施氮处理后, 中氮处理(120 kg N/(hm2 ·a))和高氮处理(240 kg N/(hm2 ·a))均明显促进杉木胸径的生长.在鼎湖山的研究也发现, 中氮处理(100 kg N/(hm2 ·a))使马尾松林胸径生长增加 77.8%[32 ] , 高氮处理则将逐渐对乔木幼苗生长产生负面效应[22 ] .通过对米槠的季节性生长分析发现, 氮添加的生长促进作用主要在水热条件较好的湿润季节产生, 这可能是由于研究地区夏季高温多雨, 土壤氮的周转速率增强, 提高了植物生长过程中氮素的可利用性[34 ] , 当氮素为限制因子时, 额外的氮输入可通过增加土壤和凋落物有效氮[35 ] 、改变基质的 C/N 比[6 ] 及改变土壤微生物组成[36 ] 等方式, 短期提高土壤的氮矿化速率, 进一步提高土壤氮素的可利用性. ...
Microbial carbon and nitrogen dynamics in coniferous forest floor material collected along a European nitrogen deposition gradient.
1
1998
... 氮添加在总体水平上增加了米槠的全年胸径相对生长速率, 说明研究区内氮素是米槠生长的限制因素.研究区内米槠在不同氮添加水平下的响应不同, 除高氮处理外, 低氮和中氮处理均显著提高了米槠的全年径向生长速率.这与我国目前在热带及亚热带森林生态系统的多数氮沉降试验结果一致, 氮沉降会促进受氮限制的年轻森林中树木的生 长[22 ,31 -33 ] , 树木对氮沉降的响应也受树木种类、研究地点和施肥年限等因素的影响.樊后保等[31 ] 对亚热带杉木的生长研究结果表明, 经过 3 年的施氮处理后, 中氮处理(120 kg N/(hm2 ·a))和高氮处理(240 kg N/(hm2 ·a))均明显促进杉木胸径的生长.在鼎湖山的研究也发现, 中氮处理(100 kg N/(hm2 ·a))使马尾松林胸径生长增加 77.8%[32 ] , 高氮处理则将逐渐对乔木幼苗生长产生负面效应[22 ] .通过对米槠的季节性生长分析发现, 氮添加的生长促进作用主要在水热条件较好的湿润季节产生, 这可能是由于研究地区夏季高温多雨, 土壤氮的周转速率增强, 提高了植物生长过程中氮素的可利用性[34 ] , 当氮素为限制因子时, 额外的氮输入可通过增加土壤和凋落物有效氮[35 ] 、改变基质的 C/N 比[6 ] 及改变土壤微生物组成[36 ] 等方式, 短期提高土壤的氮矿化速率, 进一步提高土壤氮素的可利用性. ...
氮添加对樟树林生长的影响
1
2013
... 以往的研究表明, 不同的树木径级对氮添加表现出不同的响应.郑威等[37 ] 对湖南樟树人工林的施肥实验表明, 施氮能够明显提高8 cm和 30 cm径阶的林木生长率.本研究样地内物种组成丰富, 米槠作为样地所在群落的优势种, 树木径级的不同可通过影响树木对光竞争、养分竞争及维持呼吸等方面来影响米槠的径向生长[38 -39 ] .低径级米槠对低氮和中氮处理响应强烈, 可能是由于低径级米槠生长代谢较为活跃, 冠层的快速增长导致对氮素需求较大.中等径级的米槠为样地中的优势个体, 除中氮处理外, 其他氮浓度处理下胸径相对生长率均随胸径增粗而提高, 但由于部分资源要分配给生殖生长以及根系和树干呼吸的增强[40 ] , 高氮处理使其相对生长速率显著提升.随着径级继续增大, 米槠多为老龄个体, 生长活力下降[41 ] , 光合作用产物主要用于维持性呼吸代谢[39 ] , 因而其径向生长对施氮可能无明显响应. ...
Above- and belowground biomass and primary productivity of a Larix gmelinii stand near Tura, central Siberia.
1
1999
... 以往的研究表明, 不同的树木径级对氮添加表现出不同的响应.郑威等[37 ] 对湖南樟树人工林的施肥实验表明, 施氮能够明显提高8 cm和 30 cm径阶的林木生长率.本研究样地内物种组成丰富, 米槠作为样地所在群落的优势种, 树木径级的不同可通过影响树木对光竞争、养分竞争及维持呼吸等方面来影响米槠的径向生长[38 -39 ] .低径级米槠对低氮和中氮处理响应强烈, 可能是由于低径级米槠生长代谢较为活跃, 冠层的快速增长导致对氮素需求较大.中等径级的米槠为样地中的优势个体, 除中氮处理外, 其他氮浓度处理下胸径相对生长率均随胸径增粗而提高, 但由于部分资源要分配给生殖生长以及根系和树干呼吸的增强[40 ] , 高氮处理使其相对生长速率显著提升.随着径级继续增大, 米槠多为老龄个体, 生长活力下降[41 ] , 光合作用产物主要用于维持性呼吸代谢[39 ] , 因而其径向生长对施氮可能无明显响应. ...
Declining forest productivity in aging forest stands: a modeling analy-sis of alternative hypotheses.
2
1996
... 以往的研究表明, 不同的树木径级对氮添加表现出不同的响应.郑威等[37 ] 对湖南樟树人工林的施肥实验表明, 施氮能够明显提高8 cm和 30 cm径阶的林木生长率.本研究样地内物种组成丰富, 米槠作为样地所在群落的优势种, 树木径级的不同可通过影响树木对光竞争、养分竞争及维持呼吸等方面来影响米槠的径向生长[38 -39 ] .低径级米槠对低氮和中氮处理响应强烈, 可能是由于低径级米槠生长代谢较为活跃, 冠层的快速增长导致对氮素需求较大.中等径级的米槠为样地中的优势个体, 除中氮处理外, 其他氮浓度处理下胸径相对生长率均随胸径增粗而提高, 但由于部分资源要分配给生殖生长以及根系和树干呼吸的增强[40 ] , 高氮处理使其相对生长速率显著提升.随着径级继续增大, 米槠多为老龄个体, 生长活力下降[41 ] , 光合作用产物主要用于维持性呼吸代谢[39 ] , 因而其径向生长对施氮可能无明显响应. ...
... [39 ], 因而其径向生长对施氮可能无明显响应. ...
Hydraulic limits to tree height and tree growth.
1
1997
... 以往的研究表明, 不同的树木径级对氮添加表现出不同的响应.郑威等[37 ] 对湖南樟树人工林的施肥实验表明, 施氮能够明显提高8 cm和 30 cm径阶的林木生长率.本研究样地内物种组成丰富, 米槠作为样地所在群落的优势种, 树木径级的不同可通过影响树木对光竞争、养分竞争及维持呼吸等方面来影响米槠的径向生长[38 -39 ] .低径级米槠对低氮和中氮处理响应强烈, 可能是由于低径级米槠生长代谢较为活跃, 冠层的快速增长导致对氮素需求较大.中等径级的米槠为样地中的优势个体, 除中氮处理外, 其他氮浓度处理下胸径相对生长率均随胸径增粗而提高, 但由于部分资源要分配给生殖生长以及根系和树干呼吸的增强[40 ] , 高氮处理使其相对生长速率显著提升.随着径级继续增大, 米槠多为老龄个体, 生长活力下降[41 ] , 光合作用产物主要用于维持性呼吸代谢[39 ] , 因而其径向生长对施氮可能无明显响应. ...
Functional traits shape ontogenetic growth trajectories of rain forest tree species.
1
2011
... 以往的研究表明, 不同的树木径级对氮添加表现出不同的响应.郑威等[37 ] 对湖南樟树人工林的施肥实验表明, 施氮能够明显提高8 cm和 30 cm径阶的林木生长率.本研究样地内物种组成丰富, 米槠作为样地所在群落的优势种, 树木径级的不同可通过影响树木对光竞争、养分竞争及维持呼吸等方面来影响米槠的径向生长[38 -39 ] .低径级米槠对低氮和中氮处理响应强烈, 可能是由于低径级米槠生长代谢较为活跃, 冠层的快速增长导致对氮素需求较大.中等径级的米槠为样地中的优势个体, 除中氮处理外, 其他氮浓度处理下胸径相对生长率均随胸径增粗而提高, 但由于部分资源要分配给生殖生长以及根系和树干呼吸的增强[40 ] , 高氮处理使其相对生长速率显著提升.随着径级继续增大, 米槠多为老龄个体, 生长活力下降[41 ] , 光合作用产物主要用于维持性呼吸代谢[39 ] , 因而其径向生长对施氮可能无明显响应. ...



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}