北京大学学报(自然科学版)
2017
,
53
(3):
588-596
https://doi.org/10.13209/j.0479-8023.2017.020
Orginal Article
持久性有机污染物(POPs)在水生生态系统中的环境行为
冯秋园 1 吴桐 1 万祎 2 刘学勤 3 刘永 1 ,
1. 北京大学环境科学与工程学院, 水沙科学教育部重点实验室, 北京100871 2. 北京大学城市与环境学院, 地表过程与模拟部重点实验室, 北京100871 3. 中国科学院水生生物研究所, 武汉 430072
Environmental Behavior of Persistent Organic Pollutants (POPs) in Aquatic Ecosystem
Qiuyuan FENG 1 Tong WU 1 Yi WAN 2 Xueqin LIU 3 Yong LIU 1 ,
1. College of Environmental Science and Engineering, The Key Laboratory of Water and Sediment Sciences (MOE), Peking University, Beijing 100871 2. College of Urban and Environmental Science, MOE Laboratory for Earth Surface Process, Peking University, Beijing 100871 3. Institute of Hydrobiology Chinese Academy of Science, Wuhan 430072;
通讯作者:
† 通信作者, E-mail: yongliu@pku.edu.cn
收稿日期:
2015-11-6
修回日期:
2015-12-24
网络出版日期:
2017-02-12
版权声明:
2017 《北京大学学报(自然科学版)》编辑部 《北京大学学报(自然科学版)》编辑部 所有
基金资助:
国家重点基础研究发展计划(2015CB458900)资助
摘要
作者综述并分析了持久性有机污染物(POPs)在水生生态系统的环境介质、生物个体中的环境行为及其主要影响因素, 主要结论如下。1) 风速、温度、生产力、溶氧、颗粒物的成分、粒径等气象和理化因素通过改变 POPs 在水-气界面及水柱-沉积物界面的传递以及 POPs 的降解、沉降、再悬浮、吸附、解吸等过程, 影响 POPs 在水生生态系统环境介质中的环境行为。2) POPs 的种类、分子结构、大小、亲脂性等理化性质会不同程度地影响其在生物体内的被动扩散和代谢分解, 影响生物富集性大小。在不同的条件下, 起主导作用的因素不同, 且各因素间互相影响, 起到协同或拮抗作用。3) 生物个体的生长率越大, 生物转化越强, 生物富集程度越小, 脂质含量越高, 生物富集能力越强。4) 雌性个体的生物富集能力弱于雄性, 不同条件下体型对生物富集的影响不同。当前有关 POPs 在环境介质、生物个体环境行为中的研究大多是独立开展的, 缺少对其互相联系与影响的综合分析, 也缺少在不同时空尺度上动态变化的研究。食物网是 POPs 传递的一个重要途径, 但是由于其结构的复杂多变, 是研究的重点和难点, 未来需要更深入的探索, 特别需要加强对底栖食物网、浮游-底栖耦合食物网及微食物网在POPs传递中的作用的研究。
关键词:
持久性有机污染物(POPs)
;
水生生态系统
;
环境介质
;
生物个体
;
环境行为
Abstract
This paper reviewed the environmental behaviors of POPs (persistent organic pollutants) in water column, sediment and living organism in aquatic ecosystem. The main results were presented as below. 1) Meteorological factors and physicochemical properties could determine the transmission of POPs in the interface of air-water and water column-sediment, including wind speed, temperature, productivity, dissolved oxygen (DO), particle size and component of sediment. The degradation, sedimentation, resuspension, adsorption and desorption of POPs would also be affected. 2) The categories, molecular structures, molecular size and lipophicity were closely related with the passive diffusion and degradation of POPs. Under different condition, the dominant influence factors would change. 3) Larger growth and biotransformation rate would cause smaller bioaccumulation, while higher lipid content had the opposite effects. 4) Comparing to male individuals, the female ones always had stronger bioaccumulation of POPs. The body size had different effects on bioaccumulation among various species and POPs. The analysis also found that previous studies were always conducted separately and lack of comprehen-sive investigation of POPs behavior in environmental medium and individuals, and the dynamic changes at different temporal and spatial scales were still not clear. Food web is an improtant transfer path for POPs, and in the future, more efforts should be put to explore and understand the complex behaviors of POPs in aquatic food web, especially the benthic, coupled benthic-pelagic and microbial food web.
Keywords:
persistent organic pollutants (POPs)
;
aquatic system
;
environmental medium
;
individuals
;
environmental behavior
PDF (502KB)
元数据
多维度评价
相关文章
收藏文章
本文引用格式
导出
EndNote
Ris
Bibtex
冯秋园 , 吴桐 , 万祎 , 刘学勤 , 刘永 . 持久性有机污染物(POPs)在水生生态系统中的环境行为 [J]. 北京大学学报(自然科学版) https://doi.org/10.13209/j.0479-8023.2017.020
Qiuyuan FENG , Tong WU , Yi WAN , Xueqin LIU , Yong LIU . Environmental Behavior of Persistent Organic Pollutants (POPs) in Aquatic Ecosystem [J]. Acta Scientiarum Naturalium Universitatis Pekinensis https://doi.org/10.13209/j.0479-8023.2017.020
持久性有机污染物(POPs)具有持久性、生物蓄积性、高毒性、半挥发性和远距离传输的特点, 会对人类健康和生态安全产生严重危害。2001 年, 全球120多个国家签署了持久性有机污染物的斯德哥尔摩公约, 致力于削减 POPs 污染物。POPs 通过大气干湿沉降、地面径流和水-气界面直接交换进入水生生态系统, 使其成为 POPs在生态圈中传递的一个重要的“汇”。POPs 进入水体后, 在各种物理、化学、生物作用下发生迁移和转化。生物体通过鳃、皮肤等器官的被动扩散和捕食作用吸收 POPs, 在生物体内形成生物富集效应(biocon-centration), 并沿着食物链的传递造成生物放大(biomagnification), 使得鱼类等高等水生动物体内具有相当高浓度的 POPs, 并进一步通过渔业活动对人类健康造成严重威胁。因此, 研究 POPs在环境介质以及生物个体中的环境行为, 是揭示其在水生生态系统中归趋机制的基础, 对于准确地预测POPs 生物富集性的大小, 评价暴露风险, 研究POPs 的生物、生态毒性, 制定合理的环境基准, 实施科学的管控措施具有重要意义。鉴于此, 国内外对 POPs 在水、沉积物和土壤中的污染以及在不同环境介质和生物之间的迁移转化开展了大量的研 究[1 ,2 ,3 ,4 ,5 ,6 ,7 ,8 ] 。目前国内有关水生生态系统中 POPs 的研究主要包括 POPs 在不同水生生态系统的水柱、沉积物、生物体内分布情况和浓度的研究[5 ,9 -15 ] , POPs在食物链中生物富集性的研究[1 ,2 ,3 ,4 ,5 ,6 ,7 ,8 ] , POPs的类型、性质、源解析和治理方法的研究[1 ,2 ,3 ,4 ,5 ,6 ,7 ,8 ] 这三方面。对POPs在水生生态系统环境行为及其影响因素的研究和分析相对较少。另外, 已有的有关POPs在水柱、沉积物、生物体和食物网中的研究相对独立, 缺少从生态系统的角度, 整体、全面、系统的探索 POPs 在不同相之间的环境行为、影响因素和互相联系。但实际上, POPs在水生生态系统中的环境行为是一个动态的整体过程, 在不同相之间的传递和分配是互相影响、互相联系的。
本文总结了 POPs 在水生生态系统的环境介质和生物个体中传递和转化过程与潜在机制及影响因素, 探讨 POPs 在不同相之间传递的互相联系, 为系统地研究不同水生生态系统中POPs的环境行为、生物富集、动态变化和生态毒性提供 参考。
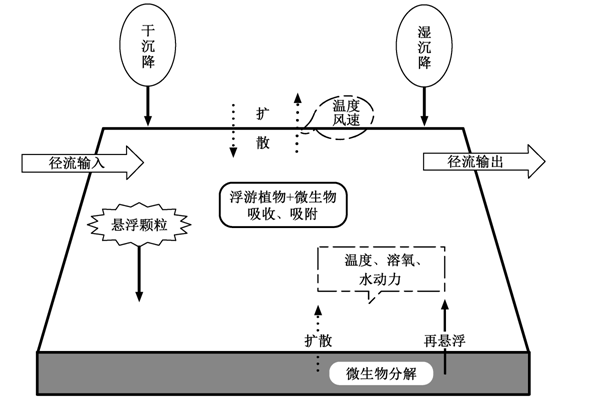
1 POPs在水柱和沉积物中的环境行为及影响因素
POPs 进入水体后, 会在各种物理、化学、生物因素影响下, 在水柱和沉积物中发生迁移和转化(图 1 )。水柱和沉积物是水生生物体生活和栖息场所, 也是其食物和能量来源。生物体通过被动扩散和摄食等活动从环境介质中吸收并富集 POPs, 并进一步沿食物网传递。因此, 研究 POPs 在水柱和沉积物等介质中的环境行为, 是研究 POPs 在水生生态系统中传递的基础, 是决定 POPs 在水生生物和食物网中生物富集和生物放大的前提。POPs 通过地表径流、大气干湿沉降、大气-水界面扩散等作用进入水体[9 ] , 其环境行为和过程受到诸多因素的影响, 如温度、风速、悬浮颗粒物、浮游植物、腐殖质含量、沉积物再悬浮和微生物等[2 ,23 -24 ] 。
1.1 POPs在水柱中的环境行为及影响因素
POPs 在水-气界面的扩散交换会受到风速和温度等因素的影响。风速通过改变大气-水表面之间的质量传质系数影响 POPs 的扩散作用; 风速越高, 温度越高, 水面 POPs 的挥发和扩散加快。水体的生产力、浮游植物和微生物的吸附以及颗粒物的沉降等也能够影响水-气界面的 POPs 转移, 进而影响POPs 在水生生态系统中的归趋。研究表明, 当水体发生富营养化时, 浮游植物的生长率和生物量会大大增加, 浮游植物的吸收会导致水柱中 PCBs 浓度降低, 打破 PCBs 在水-气界面的平衡状态, 促进PCBs从大气向水柱转移[2 ] 。当浮游植物的生物量翻倍时, 大气-水柱之间某些有机污染物的交换通量能够增加 32%~37%, 一般低氯代污染物的增加量要比高氯代污染物的增加量多。在水柱中, POPs被颗粒物(主要是浮游植物)大量吸附和吸收后, 随着颗粒物的沉降向底栖生境转移, 增加了沉积物中POPs的浓度[24 ] 。因此, 沉积物成为POPs进入水环境以后的主要蓄积场所和归宿, 沉积物中的 POPs浓度通常要比相应水柱中的浓度高几个, 甚至十几个数量级[25 ] 。此外, 腐殖质的含量和成分也是影响水柱中及生物体内 POPs 浓度的重要因素。Carter等[26 ] 发现, 腐殖质吸附 POPs 的程度受到腐殖质来源、类型、pH 值、Ca 浓度、离子化程度和腐殖质浓度大小的影响。进一步的研究表明, 随着水体中腐殖质增加, POPs 浓度会不断下降, 原因是腐殖质能够大量吸附 POPs, 降低水体以及生物体内 POPs的浓度, 且对于疏水性较强的 POPs 的影响相对更大[23 ] 。
图1 POPs在大气-水柱-沉积物之间的迁移过程
Fig. 1 Transmission of POPs in air, water column and sediment
1.2 POPs在沉积物中的环境行为及影响因素
由于沉积物的化学成分和结构形态不同, 它们与 POPs 相互作用决定了 POPs 的稳定程度、迁移速率和生物有效性的差异很大。砂质沉积物中的POPs 浓度较小, 而泥质沉积物中由于有机质含量高, 吸附能力强, POPs 的浓度较大。沉积物中有机质的组成类型也对 POPs 的吸附有很大的影响, 粒径越小的颗粒物, 其有机碳含量越高, 吸附 POPs的能力越强[27 ,28 ,29 ] 。研究发现, 沉积物中总有机碳(TOC)含量与 PAHs 含量显著相关, 沉积物的粒径与OCPs和 PCBs 的含量之间有很好的相关性[30 ] 。
沉积物中的 POPs 在一定条件下会发生各种转化, 或从颗粒物上解吸, 再次进入水柱中。微生物分解是影响 POPs 含量和分布类型的重要因素, 且在不同的环境条件下, 微生物对 POPs 的降解活性和转化产物不同。例如, 对鸭川河 3 个不同站点沉积物中的林丹和六氯苯的降解研究发现, 微生物对3个站点的六氯苯(HCBs)几乎是同时开始降解的, 但对于反-林丹(TC)和顺-林丹(CC), 不同采样点开始降解的时间有很大差异。当实验结束时, 不同种类 POPs 的降解率差别很大, 分别是 HCBs>TC> CC。同时, 微生物对POPs的降解也受到环境理化条件的影响。例如, 沉积物中的有机碳(TOC)含量越高、有机污染越严重, 微生物对 POPs 的降解速率越高[31 ] 。DDT 在厌氧条件下被微生物分解为DDD, 而在好氧条件下转为 DDE[32 ] 。透光层的颗粒物在沉降过程中, 有机质不断降解, 浓度下降, 但 POPs 的浓度却不断升高, 在颗粒物沉降到沉积物表面之后, 大部分有机质被微生物降解, 此时大部分POPs 被释放出来, 再次进入水柱中循环。除微生物外, 温度等物理条件同样也是影响沉积物中POPs 再循环的重要因素。研究发现, 随着温度的升高, 沉积物中有机氯农药的释放明显加强, 且在紊动条件下的释放速率和强度远远大于静止条件 下[25 ] 。沉积物中的POPs 除通过解吸释放到水柱中外, 还可以通过沉积物的再悬浮进入水柱中。湖泊在秋季分层结束时, 沉积物中的 POPs 会随着颗粒物的再悬浮再次进入水柱中。研究证实, 现场调查到的水柱中POPs沉降通量会比大气沉降通量高1~2 个数量级, 就是由沉积物的再悬浮造成的[33 ] 。
在水生生态系统中, 风速、温度等气象因素和水动力条件会影响 POPs 在水-气界面以及水柱-沉积物中的迁移, 生产力、浮游生物、微生物、悬浮颗粒物、腐殖质等理化和生物因素会通过吸附、吸收、解吸、沉降和再悬浮作用影响水柱中溶解态POPs 的浓度, 并进一步影响生物体中 POPs 的浓度。沉积物中微生物的代谢和转化是 POPs 从水生生态系统中脱离的一个重要途径, 会受温度、溶氧等因素的影响。
分析 POPs 在环境介质中的传递和转化, 是研究 POPs 在水生生态系统中环境行为的基础和前提。目前, 有关 POPs 在水柱和沉积物中分布和污染水平的研究较多, 对 POPs 在水-沉积物介质之间以及大气-水-沉积物介质之间的传递、转化动态行为及影响因素的深入、系统分析相对匮乏。另外, POPs 在环境介质中传递的影响因素纷繁复杂, 互相联系, 但目前的研究往往只关注其中的几个主要因素, 缺少对这些因素的综合考虑和动态变化研究。环境介质的理化性质受到生物群落和食物网结构变化的反馈作用, 因此, 单纯从环境介质的角度分析 POPs 的行为, 很难准确把握其潜在的归趋机制。
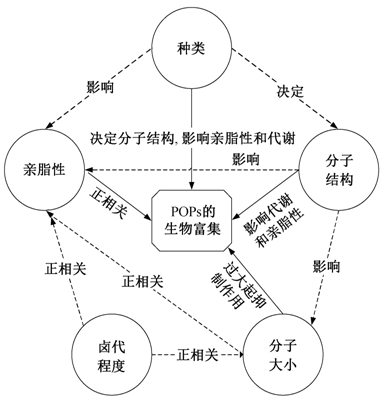
图2 POPs的理化性质对生物富集性的影响
Fig. 2 Influence of phy-chemical properties of POPs on bioaccumulation
2 POPs 在生物个体中的环境行为及影响因素
POPs 在进入水生生态系统以后, 会通过生物体的皮肤、鳃等器官的被动扩散和捕食作用进入到生物体内, POPs 在环境介质和生物体之间的分配和传递作用受诸多因素的影响: 1) POPs 理化性质; 2) 生物体生理性质。
2.1 POPs理化性质的影响
POPs 的理化性质主要包括种类、分子大小、分子结构、亲脂性、卤代程度等, 但它们之间互相关联。POPs 的苯环数量越多, 卤代程度越高, 分子越大, 亲脂性越强。在不同情况下, 这几方面因素在 POPs 生物富集过程起的作用不同, 有时是协同作用, 有时作用是相反的[3 ,34 -35 ] 。例如, 氯代程度越高, 分子越大, 亲脂性越强, 因此生物富集性会越高; 但氯代程度越大, 分子也会变大, 由于阻碍作用又会降低生物富集性。因此, 在研究 POPs 的生物富集性时, 尤其是同一类 POPs 的各种同系物的生物富集性时, 需同时评价这几类因素的影响(图2 )。
1) POPs 的亲脂性和卤代程度会影响生物富集作用及变化。Nizzetto 等[36 ] 在意大利南部的马久里湖发现, 春季水华爆发前后, 对于 lgK ow >6.7 (K ow 为辛醇-水分配系数)的 PCBs 来说, 水柱中悬浮颗粒的吸附量占绝大部分, 约为 50%~75%; 对于lgK ow <6.7 的 PCBs 来说, 水柱中溶解态的 PCBs 占优势, 为 5%~35%。在浮游动物体内, 低氯代 PCBs (lgK ow <6.45)的生物富集因子(BAFs)的最大值发生在水华爆发高峰期, 高氯代PCBs (lgK ow >6.45)的生物富集因子(BAFs)最大值发生在水华爆发之前。
2) POPs 的种类、亲脂性和结构会影响其生物集性及其在生物体内的生物转化和代谢分解。Mizukawa 等[3 ] 分析日本东京湾中双壳类、螃蟹、鱼、海水以及沉积物中所含的 20 种 PBDEs 和PCBs 的生物富集性, 发现 PBDEs 和 PCBs 的生物浓缩因子(BCFs)随着 K ow 的增加而增加, 但是当lgK ow >7以后, BCFs 随着 K ow 的增加而下降。PCBs和大多数的 PBDEs 同系物存在生物放大作用, 但另外几种 PBDEs 同系物不存在生物放大作用, 其原因可能是在生物体内被生物转化或代谢分解了。Hoekstra等[37 ] 研究了几类有机污染物在阿拉斯加和加拿大极地地区的桡足类生物体内的生物富集性, 其大小顺序为: 毒杀芬>PCBs>HCHs>DDT> CHLOR >CIBz。毒杀芬和 PCBs 生物富集性之所以较大, 是因为生物体内细胞色素 P-4502B 同工酶对这两类物质的生物转化量较少。另有研究发现, 疏水性相同的PAHs 的生物富集性小于 PCBs, 因为PAHs 在甲壳类和其他无脊椎动物体内较容易被代谢而排出[35 ,38 ] 。
3) 分子结构和卤代程度会影响 POPs 的生物富集性。例如, 7氯代和8氯代二口恶英和呋喃的营养级放大系数非常低, 原因是这些物质的分子较大, 限制了它们通过细胞膜进入细胞, 并且生物体可以通过排泄作用将它们排出体外[35 ,39 ] 。Opperhuizen等[40 ] 也认为疏水性很强的PCDDs和PCDFs在生物体内的生物富集性不高是由于氯代程度高和分子结构较大, 阻止其扩散和吸收, 降低了生物富集性。
4) POPs 的分子量和亲脂性影响生物富集性。Khairy 等[35 ] 研究了几种有机污染物在河口食物网的分布, 结果显示, 小分子量 PAHs 的脂溶性较低(lgK lip-w <4.1), 生物富集因子(lgBAFs)随着脂溶性(lgK lip-w )的增加而线性增长; 分子量增大, 脂溶性增加(4.1<lgK lip-w <5.4), 生物富集因子(lgBAFs)的增加变缓。原因是分子量较低时, PAHs 在水中的溶解度较大, 且在生物体内的生物转化率较低, 吸收速率远高于清除速率, 因此有明显的生物富集现象。而当分子量增大时, PAHs 的生物转化率提高, 生物富集系数降低。
一般情况下, POPs的疏水性(K ow )是预测其在生物体或食物网内生物富集、生物放大的重要指标和因素, 但实际上, 即使POPs 的疏水性(K ow )相同, 它们在环境和生物体内的分布模式也是不同的, 与它们的种类、结构和分子大小等具有很大的关系。POPs 的这些理化性质之间是互相联系、互相影响的, 因此, 在研究 POPs 在环境或生物体内的分布或是生物富集效应时, 需要综合考虑并识别上述不同因素的影响(图2 )。
2.2 生物个体因素的影响
研究证明, 生物个体的生理特征等因素(如: 生长率、性别、脂含量、生物转化和体型等)会影响POPs的生物富集性[41 ,42 ] (表1 )。
生长率的提高会降低生物体内POPs 浓度。在夏季富营养化的湖泊中, 浮游植物生长最快, 生物量增加, 此时体内 POPs 浓度较低, 尤其是高氯代PCBs 等疏水性较强的化合物。原因在于: 1) 浮游植物大量生长、生物量增大, 形成稀释效应; 2) 浮游植物对 POPs 的大量吸收, 打破了水-气界面的平衡分配, 降低了水柱中溶解态POPs的浓度; 3) 浮游植物对 POPs 的吸收分为两个主要的步骤, 首先是POPs 被吸附在浮游植物的表面(较快), 之后 POPs从细胞的表面转移到细胞内部(较慢), 分子越大, 脂溶性越强, 转移越慢。当浮游植物生长率较快时, POPs 在浮游植物和水柱之间分配失衡。因此浮游植物的生长率越快, 生物量越大, POPs 的脂溶性越强, 生长稀释效应就越大。不仅浮游植物如此, 鱼类等其他生物当生长率大大提高的时候, 其体内的POPs浓度也会下降[23 ] 。
雄性生物体内的 POPs 浓度高于雌性, 这是因为雌性生物会通过产卵降低体内 POPs 的浓度。研究发现, 雌性梭子鱼体内的 POPs 浓度随着年龄、体重和体型的增大而线性下降, 这是由雌鱼的季节性产卵造成的, 卵中含有的脂质含量比肌肉高10 倍, 其污染物的浓度也相应地高 10 倍左右, 而雄性通过性器官排泄的污染物较少[23 ] 。此外, McManus 等[43 ] 发现, 雌性桡足类动物在产卵过程中, 体内的 PCBs 浓度也下降较快, 同样是由于很大一部分 PCBs 转移到富含脂肪的卵上。Lotufo[44 ] 将小型的底栖桡足类暴露在荧蒽中, 发现有 50%的化合物留在卵中, 通过产卵排出体外。
生物体的脂质含量与其体内的 POPs浓度具有显著的正相关关系。Kidd 等[45 ] 发现 DDT 和 DDE的浓度与生物体脂质含量显著正相关, 且浮游生物要比相同营养级的底栖生物体内的 DDT 浓度高, 推测原因是前者的脂质含量比后者高。另外, 在寡营养湖泊中, 浮游生物、鱼类等生物体内的 POPs浓度比富营养条件下高很多, 除上面提到的生物稀释等因素以外, 寡营养条件下生物体内脂质含量更高也是一个重要原因[33 ] 。
POPs 在生物体内的生物转化能够降低其生物富集性。鱼类和很多的水生生物能够降解代谢不同种类的 POPs 物质。例如, 一种吡啶酯的实际生物富集系数是 5, 但根据辛醇-水分配系数(K ow )估算得到的生物富集系数为 678, 原因就是生物转化降低了污染物的浓度[39 ] 。另外, 前面也提到, 相同脂溶性(K ow )的 PAHs 的生物富集系数低于 PCBs, 原因就是生物体更容易分解代谢PAHs[35 ,38 ] 。
POPs 在生物体内的生物富集与体型大小有很大的关系, 但对于不同物种、不同种类的POPs, 其相关关系是不同的。Davies 等[46 ] 认为很多POPs生物富集性的变化和大小主要是由体型造成的。有研究发现, 鱼类体型增大, 脂质含量会增加, 但是其体内邻苯二甲酸酯的生物浓缩因子(BCFs)却下降为原来的 1/25[47 ] 。另有研究发现, 鱼类单位生物量含有的DDT含量同样也随着体型的增加而下降[48 ] 。这是由生物体的异速增长和生理过程变化造成的, 因为体型较大的鱼体对有机污染物的吸收变慢, 对污染物的储存能力下降[49 ] 。Harding 等[50 ] 的研究结果与此相反, 他们发现浮游生物体内单位脂质的PCBs含量不会随着体型大小的变化而变化, 鱼类单位脂质中 PCBs 含量会随着体型的增大而增大。Kucklick 等[51 ] 在苏必利尔湖的研究中发现, 随着体型的增大, 胡瓜鱼单位鲜重的 PCBs 含量从体型最小时的 21 ng/g 增长到体长为 150~199 mm 时的 53 ng/g, 但单位脂质的 PCBs 含量几乎不变, 他们认为污染物浓度随着体型增大而增大是由脂质含量随着体型增大而增加引起的。综上可知, 由于生物体的生物代谢、脂质含量等原因, 使得体型大小在POPs生物富集方面所起的作用可能是不同的。
生物个体的 POPs 生物富集性研究一直是POPs研究的重点之一, 是研究 POPs 沿食物链放大的基础, 也是研究 POPs 生物、生态毒性的前提。研究者通常从 POPs 的理化性质和生物体生理性质两个角度来分析 POPs 生物富集性的变化, 其中POPs 的疏水性(K ow )和生物个体的脂质含量是最常用的两个因素。但是, 这两个因素无法解释 POPs生物富集性的实际变化, 需要从 POPs 的其他理化性质和生物体生理性质方面综合考虑。在一定的条件下, 控制 POPs 生物富集的主导因素是变化的。另外, POPs的各理化性质和生物体的各生理性质是互相联系、互相影响的, 彼此之间起协同或拮抗作用。因此, 单纯地从某几个因素来分别探讨 POPs的生物富集性大小, 往往难以发现其规律性和动态变化。另外, 从生物体个体角度来分析和解释POPs 生物富集性的大小, 难以反映种群和群落水平上 POPs 的传递、迁移和变化, 也难以评估整个生物群落的 POPs 暴露风险。因此, 未来对个体生物POPs生物富集性的大小的研究需要进一步与种群、群落的结构特征和动态变化相结合, 同时考虑环境介质的理化性质变化对 POPs 的迁移变化和群落结构的影响。
3 结论与展望
3.1 研究结论
1) POPs在水柱和沉积物中的浓度、分布和迁移转化的研究开展较多, 是分析水生生态系统POPs污染水平的初步探索, 是研究POPs生物富集性及沿食物链生物传递的前提。研究发现, 气象和理化因素通过改变 POPs 在水-气界面及水柱-沉积物界面的迁移转化、降解、沉降、再悬浮、吸附、解吸等过程, 影响 POPs 在水生生态系统的环境介质中的环境行为。
2) 个体层面的 POPs 分配和转化是识别 POPs生物富集效应的基础, 是研究 POPs 生物、生态毒性的前提, 也是研究 POPs 在食物网和生态系统中传递的关键环节。已有研究主要从 POPs 的理化性质和生物体的生理性质两个角度分析 POPs 在生物个体中生物富集性大小, 其中, POPs的疏水性(K ow )和生物体的脂质含量是解释生物富集性最常用的两个指标。亲脂性越强, 脂质含量越高, POPs在生物个体中的生物富集性越强。但是, 在一定条件下, POPs 的分子结构、大小、卤代等其他理化性质和生物体的生长率、代谢转化水平以及性别等其他生理因素占主导作用, 它们通过影响生物体对 POPs的分解代谢能力、吸收程度和排泄水平, 改变POPs 的亲脂性和脂质含量与生物富集性之间的相关关系。并且, POPs的各理化性质以及生物体的各生理性质间互相影响, 因此在研究 POPs 在个体生物中的富集性变化时, 需要在识别主要影响因素的同时, 综合考虑其他影响因素的互相作用。
3.2 研究展望
1) POPs 在水生生态系统环境介质中的分布和迁移转化的研究最为广泛, 但是, 目前往往将POPs在水柱和沉积物中环境行为的研究割裂开来, 缺乏对 POPs 在不同介质之间传递行为及动态变化的研究, 缺乏对其影响因素深入、全面的探讨, 对不同传递行为及影响因素之间的互相联系关注不够。另外, 环境介质的理化性质同时受到生物群落和食物网结构变化的反馈作用, 单纯从环境介质的角度分析 POPs 的行为, 很难准确地把握其潜在的归趋机制, 因此需要从生态系统的角度开展系统研究。
2) 目前有关 POPs 生物富集性的研究通常从POPs的理化性质和生物体的生理性质两个角度来进行分析, 其中 POPs 的疏水性(K ow )和生物个体的脂质含量关注的重点。但是, 影响 POPs 富集性的主导因素会发生变化, 且各因素之间互相联系, 互相影响, 起协同或拮抗作用。因此, 未来的研究需要综合考虑多个因素来探讨 POPs 的生物富集性和动态变化规律。另外, 个体生物的 POPs 富集性研究难以反映种群和群落水平上 POPs 的迁移、转化和暴露风险, 因此, 未来的研究需要结合种群、群落的结构特征和动态变化, 同时考虑环境介质理化性质的改变对 POPs 的迁移变化和群落结构的 影响。
3) 食物网是生态系统结构和功能的综合体现, 是POPs 在水生生态系统中传递的重要途径。由于食物网结构复杂多变, 是研究的难点。其中, 底层食物网是 POPs 进入食物网的基础, 也是能量传递和物质流动的起点, 但由于其体型小, 使得 POPs在底层食物网的研究受到限制, 未来应给予更多的关注。底栖及底栖-浮游耦合食物在生态系统功能方面的作用逐渐受到人们的重视, 因此 POPs 在底栖及底栖-浮游耦合食物网中的环境行为研究已成为热点方向。微食物网是食物网的关键一环, 但目前对此方面的了解甚少, 未来应加强对 POPs 在微食物网中环境行为的研究, 对于完善 POPs 的归趋机制具有不可或缺的作用。
The authors have declared that no competing interests exist.
参考文献
文献选项
[1]
Paterson M J Muir D C G, Rosenberg B, et al. Does lake size affect concentrations of atmospherically derived polychlorinated biphenyls in water, sediment, zooplankton, and fish?
. Canadian Journal of Fisheries and Aquatic Sciences , 1998 , 55 : 544 -553
[本文引用: 3]
[2]
Dachs J Eisenreich S J Hoff R M Influence of eutrophication on air-water exchange, vertical fluxes, and phytoplankton concentrations of persistent orga-nic pollutants. Environmental Science and Techno-logy , 2000 , 34 (6 ): 1095 -1102
[本文引用: 5]
[3]
Mizukawa K Takadab H. Takeuchi I et al .Biocon-centration and biomagnification of polybrominated-diphenyl ethers (PBDEs) through lower-trophic-level coastal marine food web
. Marine Pollution Bulletin , 2009 , 58 (8 ): 1217 -1224
[本文引用: 5]
[4]
Zhang P Song J M Yuan H M Persistent organic pollutant residues in the sediments and mollusks from the Bohai Sea coastal areas, North China: an over-view
. Environment International , 2009 , 35 (3 ): 632 -646
[本文引用: 3]
[5]
刘远 . 持久性有机污染物在渤海沉积物中的分布规律与来源解析[D]
. 大连: 大连海事大学 , 2010
[本文引用: 4]
[6]
Liu W X He W Qin N et al .Residues, distributions, sources, and ecological risks of OCPs in the water from Lake Chaohu, China
. The Scientific World Journal , 2012 , 2012 : 1 -16
[本文引用: 3]
[7]
Xu F L Kong X Z He W et al .Distributions, sources, and ecological risks of hexachlorocy-clohexanes in the sediments from Haihe Plain, Northern China
. Environmental Science and Pollution Research , 2013 , 20 (4 ): 2009 -2019
[本文引用: 3]
[8]
Roots O Lukki T Přibylová P et al .Measurements of persistent organic pollutants in Estonian ambient air (1990-2013)
. Proceedings of the Estonian Academy of Sciences , 2015 , 64 (2 ): 184 -199
[本文引用: 3]
[9]
胡晴晖 . 湄洲湾水体中持久性有机污染物的污染特征与风险评估
. 中国环境科学 , 2014 , 34 (10 ): 2536 -2544
[本文引用: 2]
[10]
张宗雁 . 持久性有机污染物在东海泥质区的分布与沉积记录[D]
. 青岛: 中国海洋大学 , 2005
[11]
甘志芬 . 渤海湾西岸典型持久性有机污染物的环境行为初步研究[D]
. 保定: 河北大学 , 2010
[12]
姚威风 . 持久性有机污染物在贡湖沉积物和水体中的分布特征及源解析[D]
. 长春: 吉林农业大学 , 2010
[13]
Gao J M Hu J Y Wan Y et al .Butyltin compounds distribution in the coastal waters of Bohai Bay, People’s Republic of China. Bulletin of Environm-ental Contamination and
Toxicology , 2004 , 72 : 945 -953
[14]
Wan Y Hu J Y Liu J L et al .Fate of DDT-related compounds in Bohai Bay and its adjacent Haihe Basin, north China
. Marine Pollution Bulletin , 2005 , 50 : 439 -445
[15]
戴国华 . 影响沉积物-水界面持久性有机污染物迁移行为的因素研究
. 环境化学 , 2011 , 30 (1 ): 224 -230
[本文引用: 1]
[16]
张桂斋 . 两类持久性有机污染物和重金属在南四湖食物链中的分布和生物积累[D]
. 济南: 山东大学 , 2014
[17]
Wan Y Hu J Y Yang M et al .Characterization of trophic transfer for PCDD/Fs and coplanar PCBs in the marine food web of Bohai Bay, North China
. Environmental Science & Technology , 2005 , 39 : 2417 -2545
[18]
Hu J Y Zhen H J, Wan Y, et al. Trophic magnification of triphenyltin in marine food web: comparison to tributyltin
. Environmental Science & Technology , 2006 , 40 : 3142 -3147
[19]
李雪梅 , 张庆华 , 甘一萍 , 等 . 持久性有机污染物在食物链中积累与放大研究进展
. 应用与环境生物学报 , 2008 , 13 (6 ): 901 -905
[20]
韦朝海 , 长小璇 , 任源 , 等 . 持久性有机污染物的水污染控制: 吸附富集、生物降解与过程分析
. 环境化学 , 2011 , 30 (1 ): 300 -309
[21]
黄栩 , 骆苑蓉 , 胡忠 , 等 . 持久性有机污染物(POPs)生物修复研究进展
. 环境科学学报 , 2006 , 26 (3 ): 353 -361
[22]
员晓燕 , 杨玉义 , 李庄孝 , 等 . 中国淡水环境中典型持久性有机污染物(POPs)的污染现状与分布特征
. 环境化学 , 2013 , 32 (11 ): 2072 -2081
[23]
Larsson P Collvin L Okla L et al .Lake productivity and water chemistry as governors of the uptake of POPs in fish
. Environmental Science and Technology , 1992 , 26 (2 ): 346 -352
[本文引用: 6]
[24]
Dachs J Eisenreich S J Baker J E et al .Coupling of phytoplankton uptake and air-water exchange of persistent organic pollutants
. Environmental Science and Technology , 1999 , 33 : 3656 -3660
[本文引用: 2]
[25]
龚香宜 . 有机氯农药在湖泊水体和沉积物中的污染特征及动力学特征: 以洪湖为例[D]
. 武汉: 中国地质大学 , 2007
[本文引用: 2]
[26]
Carter C W Suffet I H Binding of DDT to dissolved humic materials. Environmental Science and Techno-logy , 1982 , 16 (11 ): 735 -740
[本文引用: 1]
[27]
Choi W W Chen K Y Associations of chlorinated hydrocarbons with fine Particles and humic substan-ces innearshore surficial sediments
. Environmental Science and Technology , 1976 , 10 (8 ): 782 -786
[本文引用: 1]
[28]
Karickhoff S W Brown D S Scott T A Sorption of hydrophobic Pollutants on nature sediments
. Water Research , 1979 , 13 : 241 -248
[本文引用: 1]
[29]
Young J L Spycher G Water dispersible soil organic mineral particles: I. Carbon and nitrogen distribution
. Soil Science Society of America Journal , 1979 , 43 : 324 -328
[本文引用: 1]
[30]
Zheng B H Zhao X R Liu L S et al .Effects of hydrodynamics on the distribution of trace POPs and macrobenthic communities in Bohai Bay. Chemos-phere , 2011 , 84 : 336 -341
[本文引用: 1]
[31]
Hirano T Ishida T Oh K et al .Biodegradation of chlordane and hexachorobenzenes in river sedimen
. Chemosphere , 2007 , 67 (3 ): 428 -434
[本文引用: 1]
[32]
Hitch R K Day H R Unusual Persistence of DDT in some Western USA soils
. Bulletin of Environmental Contamination and Toxicology , 1992 , 48 (2 ): 255 -264
[本文引用: 1]
[33]
Larsson P Andersson A Broman D et al .Persistent organic pollutants (POPs) in pelagic system
. Royal Swedish Academy of Science , 2000 , 29 (4/5 ): 202 -209
[本文引用: 3]
[34]
Ma Y G Lei Y D Xiao H et al .Critical review and recommended values for the physical-chemical pro-perty data of 15 Polycyclic Aromatic Hydrocarbons at 25°C
. Journal of Chemical and Engineering Data , 2010 , 55 (2 ): 819 -825
[本文引用: 1]
[35]
Khairy M A Weinstein M P Lohmann R Trophodyna- mic behavior of hydrophobic organic contaminants in the aquatic food web of a tidal river
. Environmental Science and Technology , 2014 , 48 (21 ): 12533 -12542
[本文引用: 6]
[36]
Nizzetto L Gioia R Li J et al .Biological pump control of the fate and distribution of hydrophic organic pollutants in water and plankton. Environ-mental Science and
Technology , 2012 , 46 : 3204 -3211
[本文引用: 1]
[37]
Hoekstra P F O’Hara T M, Teixeira C, et al. Spatial trends and bioaccumulation of organochlorine pollu-tants in marine zooplankton from the Alaskan and Canadian Arctic
. Environmental Toxicology and Chemistry , 2002 , 21 (3 ): 575 -583
[本文引用: 1]
[38]
Nfon E Cousins I T Broman D Biomagnification of organic pollutants in benthic and pelagic marine food chains from the Baltic Sea
. Science of the Total Environment , 2008 , 397 : 190 -204
[本文引用: 3]
[39]
Barron M G Bioconcentration. Environmental Scie-nce and
Technology , 1990 , 24 (11 ): 1612 -1618
[本文引用: 3]
[40]
Opperhuizen A Sijm D T H M. Bioaccumulation and biotransformation of polychlorinated Dibenzo-p-Dioxins and Dibenzofurans in fish
. Environmental Toxicology and Chemistry , 1990 , 9 (2 ): 175 -186
[本文引用: 1]
[41]
Kidd K A Hesslein R H Ross B J et al .Bio-accumulation of organochlorines through a remote freshwater food web in the Canadian Arctic
. Environmental Pollution , 1998 , 102 (1 ): 91 -103
[本文引用: 1]
[42]
Kannan K Perrotta E Thomas N J Association between perfluorinated compounds and pathological conditions in southern sea otters
. Environmental Science and Technology , 2006 , 40 (16 ): 4943 -4948
[本文引用: 1]
[43]
McManus G B Wyman K D Peterson W T et al . Factors affecting the elimination of PCBs in the marine copepod Acartiatonsa
. Estuarine Coastal and Shelf Science , 1983 , 17 (4 ): 421 -430
[本文引用: 2]
[44]
Lotufo G R Bioaccumulation of sediment-associated fluoranthene in benthic copepods: uptake, elimination and biotransformation
. Aquatic Toxicology , 1998 , 44 : 1 -15
[本文引用: 2]
[45]
Kidd K A Bootsma H A Hesslein R H et al .Biomagnification of DDT through the benthic and pelagic food webs of Lake Malawi, East Africa: importance of trophic level and carbon source
. Environmental Science and Technology , 2001 , 35 (1 ): 14 -20
[本文引用: 2]
[46]
Davies R P Dobbs A J The prediction of biocon-centration in fish
. Water Research , 1984 , 18 (10 ): 1253 -1262
[本文引用: 2]
[47]
Tarr B D Barron M G. Hayton W L. Effect of body size on the uptake and bioconcentration of di-2-ethylhexyl phthalate in rainbow trout
. Environmental Toxicology and Chemistry , 1990 , 9 (8 ): 989 -995
[本文引用: 2]
[48]
Murphy P G Murphy J V Correlations between respiration and direct uptake of DDT in the mosquito fish Gambusiaaffinis. Bulletin of Environmental Con-tamination and
Toxicology , 1971 , 6 (6 ): 581 -588
[本文引用: 2]
[49]
Adolph E F Quantitative relations in the physiologi-cal constitutions of mammals
. Science , 1949 , 109 : 579 -585
[本文引用: 2]
[50]
Harding G C LeBlanc R J, Wass W P, et al. Bioaccumulation of polychlorinated biphenyls (PCBs) in the marine pelagic food web, based on a seasonal study in the southern Gulf of St. Lawrence, 1976-1977
. Marine Chemistry , 1997 , 56 (3/4 ): 145 -179
[本文引用: 2]
[51]
Kucklick J R. Baker J E. Organochlorines in Lake Superior’s food web
. Environmental Science and Technology , 1998 , 32 (9 ): 1192 -1198
[本文引用: 2]
Muir D C G, Rosenberg B, et al. Does lake size affect concentrations of atmospherically derived polychlorinated biphenyls in water, sediment, zooplankton, and fish?
3
1998
... 持久性有机污染物(POPs)具有持久性、生物蓄积性、高毒性、半挥发性和远距离传输的特点, 会对人类健康和生态安全产生严重危害.2001 年, 全球120多个国家签署了持久性有机污染物的斯德哥尔摩公约, 致力于削减 POPs 污染物.POPs 通过大气干湿沉降、地面径流和水-气界面直接交换进入水生生态系统, 使其成为 POPs在生态圈中传递的一个重要的“汇”.POPs 进入水体后, 在各种物理、化学、生物作用下发生迁移和转化.生物体通过鳃、皮肤等器官的被动扩散和捕食作用吸收 POPs, 在生物体内形成生物富集效应(biocon-centration), 并沿着食物链的传递造成生物放大(biomagnification), 使得鱼类等高等水生动物体内具有相当高浓度的 POPs, 并进一步通过渔业活动对人类健康造成严重威胁.因此, 研究 POPs在环境介质以及生物个体中的环境行为, 是揭示其在水生生态系统中归趋机制的基础, 对于准确地预测POPs 生物富集性的大小, 评价暴露风险, 研究POPs 的生物、生态毒性, 制定合理的环境基准, 实施科学的管控措施具有重要意义.鉴于此, 国内外对 POPs 在水、沉积物和土壤中的污染以及在不同环境介质和生物之间的迁移转化开展了大量的研 究[1 ,2 ,3 ,4 ,5 ,6 ,7 ,8 ] .目前国内有关水生生态系统中 POPs 的研究主要包括 POPs 在不同水生生态系统的水柱、沉积物、生物体内分布情况和浓度的研究[5 ,9 -15 ] , POPs在食物链中生物富集性的研究[1 ,2 ,3 ,4 ,5 ,6 ,7 ,8 ] , POPs的类型、性质、源解析和治理方法的研究[1 ,2 ,3 ,4 ,5 ,6 ,7 ,8 ] 这三方面.对POPs在水生生态系统环境行为及其影响因素的研究和分析相对较少.另外, 已有的有关POPs在水柱、沉积物、生物体和食物网中的研究相对独立, 缺少从生态系统的角度, 整体、全面、系统的探索 POPs 在不同相之间的环境行为、影响因素和互相联系.但实际上, POPs在水生生态系统中的环境行为是一个动态的整体过程, 在不同相之间的传递和分配是互相影响、互相联系的. ...
... [1 ,2 ,3 ,4 ,5 ,6 ,7 ,8 ], POPs的类型、性质、源解析和治理方法的研究[1 ,2 ,3 ,4 ,5 ,6 ,7 ,8 ] 这三方面.对POPs在水生生态系统环境行为及其影响因素的研究和分析相对较少.另外, 已有的有关POPs在水柱、沉积物、生物体和食物网中的研究相对独立, 缺少从生态系统的角度, 整体、全面、系统的探索 POPs 在不同相之间的环境行为、影响因素和互相联系.但实际上, POPs在水生生态系统中的环境行为是一个动态的整体过程, 在不同相之间的传递和分配是互相影响、互相联系的. ...
... [1 ,2 ,3 ,4 ,5 ,6 ,7 ,8 ]这三方面.对POPs在水生生态系统环境行为及其影响因素的研究和分析相对较少.另外, 已有的有关POPs在水柱、沉积物、生物体和食物网中的研究相对独立, 缺少从生态系统的角度, 整体、全面、系统的探索 POPs 在不同相之间的环境行为、影响因素和互相联系.但实际上, POPs在水生生态系统中的环境行为是一个动态的整体过程, 在不同相之间的传递和分配是互相影响、互相联系的. ...
5
2000
... 持久性有机污染物(POPs)具有持久性、生物蓄积性、高毒性、半挥发性和远距离传输的特点, 会对人类健康和生态安全产生严重危害.2001 年, 全球120多个国家签署了持久性有机污染物的斯德哥尔摩公约, 致力于削减 POPs 污染物.POPs 通过大气干湿沉降、地面径流和水-气界面直接交换进入水生生态系统, 使其成为 POPs在生态圈中传递的一个重要的“汇”.POPs 进入水体后, 在各种物理、化学、生物作用下发生迁移和转化.生物体通过鳃、皮肤等器官的被动扩散和捕食作用吸收 POPs, 在生物体内形成生物富集效应(biocon-centration), 并沿着食物链的传递造成生物放大(biomagnification), 使得鱼类等高等水生动物体内具有相当高浓度的 POPs, 并进一步通过渔业活动对人类健康造成严重威胁.因此, 研究 POPs在环境介质以及生物个体中的环境行为, 是揭示其在水生生态系统中归趋机制的基础, 对于准确地预测POPs 生物富集性的大小, 评价暴露风险, 研究POPs 的生物、生态毒性, 制定合理的环境基准, 实施科学的管控措施具有重要意义.鉴于此, 国内外对 POPs 在水、沉积物和土壤中的污染以及在不同环境介质和生物之间的迁移转化开展了大量的研 究[1 ,2 ,3 ,4 ,5 ,6 ,7 ,8 ] .目前国内有关水生生态系统中 POPs 的研究主要包括 POPs 在不同水生生态系统的水柱、沉积物、生物体内分布情况和浓度的研究[5 ,9 -15 ] , POPs在食物链中生物富集性的研究[1 ,2 ,3 ,4 ,5 ,6 ,7 ,8 ] , POPs的类型、性质、源解析和治理方法的研究[1 ,2 ,3 ,4 ,5 ,6 ,7 ,8 ] 这三方面.对POPs在水生生态系统环境行为及其影响因素的研究和分析相对较少.另外, 已有的有关POPs在水柱、沉积物、生物体和食物网中的研究相对独立, 缺少从生态系统的角度, 整体、全面、系统的探索 POPs 在不同相之间的环境行为、影响因素和互相联系.但实际上, POPs在水生生态系统中的环境行为是一个动态的整体过程, 在不同相之间的传递和分配是互相影响、互相联系的. ...
... ,2 ,3 ,4 ,5 ,6 ,7 ,8 ], POPs的类型、性质、源解析和治理方法的研究[1 ,2 ,3 ,4 ,5 ,6 ,7 ,8 ] 这三方面.对POPs在水生生态系统环境行为及其影响因素的研究和分析相对较少.另外, 已有的有关POPs在水柱、沉积物、生物体和食物网中的研究相对独立, 缺少从生态系统的角度, 整体、全面、系统的探索 POPs 在不同相之间的环境行为、影响因素和互相联系.但实际上, POPs在水生生态系统中的环境行为是一个动态的整体过程, 在不同相之间的传递和分配是互相影响、互相联系的. ...
... ,2 ,3 ,4 ,5 ,6 ,7 ,8 ]这三方面.对POPs在水生生态系统环境行为及其影响因素的研究和分析相对较少.另外, 已有的有关POPs在水柱、沉积物、生物体和食物网中的研究相对独立, 缺少从生态系统的角度, 整体、全面、系统的探索 POPs 在不同相之间的环境行为、影响因素和互相联系.但实际上, POPs在水生生态系统中的环境行为是一个动态的整体过程, 在不同相之间的传递和分配是互相影响、互相联系的. ...
... POPs 进入水体后, 会在各种物理、化学、生物因素影响下, 在水柱和沉积物中发生迁移和转化(图 1 ).水柱和沉积物是水生生物体生活和栖息场所, 也是其食物和能量来源.生物体通过被动扩散和摄食等活动从环境介质中吸收并富集 POPs, 并进一步沿食物网传递.因此, 研究 POPs 在水柱和沉积物等介质中的环境行为, 是研究 POPs 在水生生态系统中传递的基础, 是决定 POPs 在水生生物和食物网中生物富集和生物放大的前提.POPs 通过地表径流、大气干湿沉降、大气-水界面扩散等作用进入水体[9 ] , 其环境行为和过程受到诸多因素的影响, 如温度、风速、悬浮颗粒物、浮游植物、腐殖质含量、沉积物再悬浮和微生物等[2 ,23 -24 ] . ...
... POPs 在水-气界面的扩散交换会受到风速和温度等因素的影响.风速通过改变大气-水表面之间的质量传质系数影响 POPs 的扩散作用; 风速越高, 温度越高, 水面 POPs 的挥发和扩散加快.水体的生产力、浮游植物和微生物的吸附以及颗粒物的沉降等也能够影响水-气界面的 POPs 转移, 进而影响POPs 在水生生态系统中的归趋.研究表明, 当水体发生富营养化时, 浮游植物的生长率和生物量会大大增加, 浮游植物的吸收会导致水柱中 PCBs 浓度降低, 打破 PCBs 在水-气界面的平衡状态, 促进PCBs从大气向水柱转移[2 ] .当浮游植物的生物量翻倍时, 大气-水柱之间某些有机污染物的交换通量能够增加 32%~37%, 一般低氯代污染物的增加量要比高氯代污染物的增加量多.在水柱中, POPs被颗粒物(主要是浮游植物)大量吸附和吸收后, 随着颗粒物的沉降向底栖生境转移, 增加了沉积物中POPs的浓度[24 ] .因此, 沉积物成为POPs进入水环境以后的主要蓄积场所和归宿, 沉积物中的 POPs浓度通常要比相应水柱中的浓度高几个, 甚至十几个数量级[25 ] .此外, 腐殖质的含量和成分也是影响水柱中及生物体内 POPs 浓度的重要因素.Carter等[26 ] 发现, 腐殖质吸附 POPs 的程度受到腐殖质来源、类型、pH 值、Ca 浓度、离子化程度和腐殖质浓度大小的影响.进一步的研究表明, 随着水体中腐殖质增加, POPs 浓度会不断下降, 原因是腐殖质能够大量吸附 POPs, 降低水体以及生物体内 POPs的浓度, 且对于疏水性较强的 POPs 的影响相对更大[23 ] . ...
Biocon-centration and biomagnification of polybrominated-diphenyl ethers (PBDEs) through lower-trophic-level coastal marine food web
5
2009
... 持久性有机污染物(POPs)具有持久性、生物蓄积性、高毒性、半挥发性和远距离传输的特点, 会对人类健康和生态安全产生严重危害.2001 年, 全球120多个国家签署了持久性有机污染物的斯德哥尔摩公约, 致力于削减 POPs 污染物.POPs 通过大气干湿沉降、地面径流和水-气界面直接交换进入水生生态系统, 使其成为 POPs在生态圈中传递的一个重要的“汇”.POPs 进入水体后, 在各种物理、化学、生物作用下发生迁移和转化.生物体通过鳃、皮肤等器官的被动扩散和捕食作用吸收 POPs, 在生物体内形成生物富集效应(biocon-centration), 并沿着食物链的传递造成生物放大(biomagnification), 使得鱼类等高等水生动物体内具有相当高浓度的 POPs, 并进一步通过渔业活动对人类健康造成严重威胁.因此, 研究 POPs在环境介质以及生物个体中的环境行为, 是揭示其在水生生态系统中归趋机制的基础, 对于准确地预测POPs 生物富集性的大小, 评价暴露风险, 研究POPs 的生物、生态毒性, 制定合理的环境基准, 实施科学的管控措施具有重要意义.鉴于此, 国内外对 POPs 在水、沉积物和土壤中的污染以及在不同环境介质和生物之间的迁移转化开展了大量的研 究[1 ,2 ,3 ,4 ,5 ,6 ,7 ,8 ] .目前国内有关水生生态系统中 POPs 的研究主要包括 POPs 在不同水生生态系统的水柱、沉积物、生物体内分布情况和浓度的研究[5 ,9 -15 ] , POPs在食物链中生物富集性的研究[1 ,2 ,3 ,4 ,5 ,6 ,7 ,8 ] , POPs的类型、性质、源解析和治理方法的研究[1 ,2 ,3 ,4 ,5 ,6 ,7 ,8 ] 这三方面.对POPs在水生生态系统环境行为及其影响因素的研究和分析相对较少.另外, 已有的有关POPs在水柱、沉积物、生物体和食物网中的研究相对独立, 缺少从生态系统的角度, 整体、全面、系统的探索 POPs 在不同相之间的环境行为、影响因素和互相联系.但实际上, POPs在水生生态系统中的环境行为是一个动态的整体过程, 在不同相之间的传递和分配是互相影响、互相联系的. ...
... ,3 ,4 ,5 ,6 ,7 ,8 ], POPs的类型、性质、源解析和治理方法的研究[1 ,2 ,3 ,4 ,5 ,6 ,7 ,8 ] 这三方面.对POPs在水生生态系统环境行为及其影响因素的研究和分析相对较少.另外, 已有的有关POPs在水柱、沉积物、生物体和食物网中的研究相对独立, 缺少从生态系统的角度, 整体、全面、系统的探索 POPs 在不同相之间的环境行为、影响因素和互相联系.但实际上, POPs在水生生态系统中的环境行为是一个动态的整体过程, 在不同相之间的传递和分配是互相影响、互相联系的. ...
... ,3 ,4 ,5 ,6 ,7 ,8 ]这三方面.对POPs在水生生态系统环境行为及其影响因素的研究和分析相对较少.另外, 已有的有关POPs在水柱、沉积物、生物体和食物网中的研究相对独立, 缺少从生态系统的角度, 整体、全面、系统的探索 POPs 在不同相之间的环境行为、影响因素和互相联系.但实际上, POPs在水生生态系统中的环境行为是一个动态的整体过程, 在不同相之间的传递和分配是互相影响、互相联系的. ...
... POPs 的理化性质主要包括种类、分子大小、分子结构、亲脂性、卤代程度等, 但它们之间互相关联.POPs 的苯环数量越多, 卤代程度越高, 分子越大, 亲脂性越强.在不同情况下, 这几方面因素在 POPs 生物富集过程起的作用不同, 有时是协同作用, 有时作用是相反的[3 ,34 -35 ] .例如, 氯代程度越高, 分子越大, 亲脂性越强, 因此生物富集性会越高; 但氯代程度越大, 分子也会变大, 由于阻碍作用又会降低生物富集性.因此, 在研究 POPs 的生物富集性时, 尤其是同一类 POPs 的各种同系物的生物富集性时, 需同时评价这几类因素的影响(图2 ). ...
... 2) POPs 的种类、亲脂性和结构会影响其生物集性及其在生物体内的生物转化和代谢分解.Mizukawa 等[3 ] 分析日本东京湾中双壳类、螃蟹、鱼、海水以及沉积物中所含的 20 种 PBDEs 和PCBs 的生物富集性, 发现 PBDEs 和 PCBs 的生物浓缩因子(BCFs)随着 K ow 的增加而增加, 但是当lgK ow >7以后, BCFs 随着 K ow 的增加而下降.PCBs和大多数的 PBDEs 同系物存在生物放大作用, 但另外几种 PBDEs 同系物不存在生物放大作用, 其原因可能是在生物体内被生物转化或代谢分解了.Hoekstra等[37 ] 研究了几类有机污染物在阿拉斯加和加拿大极地地区的桡足类生物体内的生物富集性, 其大小顺序为: 毒杀芬>PCBs>HCHs>DDT> CHLOR >CIBz.毒杀芬和 PCBs 生物富集性之所以较大, 是因为生物体内细胞色素 P-4502B 同工酶对这两类物质的生物转化量较少.另有研究发现, 疏水性相同的PAHs 的生物富集性小于 PCBs, 因为PAHs 在甲壳类和其他无脊椎动物体内较容易被代谢而排出[35 ,38 ] . ...
Persistent organic pollutant residues in the sediments and mollusks from the Bohai Sea coastal areas, North China: an over-view
3
2009
... 持久性有机污染物(POPs)具有持久性、生物蓄积性、高毒性、半挥发性和远距离传输的特点, 会对人类健康和生态安全产生严重危害.2001 年, 全球120多个国家签署了持久性有机污染物的斯德哥尔摩公约, 致力于削减 POPs 污染物.POPs 通过大气干湿沉降、地面径流和水-气界面直接交换进入水生生态系统, 使其成为 POPs在生态圈中传递的一个重要的“汇”.POPs 进入水体后, 在各种物理、化学、生物作用下发生迁移和转化.生物体通过鳃、皮肤等器官的被动扩散和捕食作用吸收 POPs, 在生物体内形成生物富集效应(biocon-centration), 并沿着食物链的传递造成生物放大(biomagnification), 使得鱼类等高等水生动物体内具有相当高浓度的 POPs, 并进一步通过渔业活动对人类健康造成严重威胁.因此, 研究 POPs在环境介质以及生物个体中的环境行为, 是揭示其在水生生态系统中归趋机制的基础, 对于准确地预测POPs 生物富集性的大小, 评价暴露风险, 研究POPs 的生物、生态毒性, 制定合理的环境基准, 实施科学的管控措施具有重要意义.鉴于此, 国内外对 POPs 在水、沉积物和土壤中的污染以及在不同环境介质和生物之间的迁移转化开展了大量的研 究[1 ,2 ,3 ,4 ,5 ,6 ,7 ,8 ] .目前国内有关水生生态系统中 POPs 的研究主要包括 POPs 在不同水生生态系统的水柱、沉积物、生物体内分布情况和浓度的研究[5 ,9 -15 ] , POPs在食物链中生物富集性的研究[1 ,2 ,3 ,4 ,5 ,6 ,7 ,8 ] , POPs的类型、性质、源解析和治理方法的研究[1 ,2 ,3 ,4 ,5 ,6 ,7 ,8 ] 这三方面.对POPs在水生生态系统环境行为及其影响因素的研究和分析相对较少.另外, 已有的有关POPs在水柱、沉积物、生物体和食物网中的研究相对独立, 缺少从生态系统的角度, 整体、全面、系统的探索 POPs 在不同相之间的环境行为、影响因素和互相联系.但实际上, POPs在水生生态系统中的环境行为是一个动态的整体过程, 在不同相之间的传递和分配是互相影响、互相联系的. ...
... ,4 ,5 ,6 ,7 ,8 ], POPs的类型、性质、源解析和治理方法的研究[1 ,2 ,3 ,4 ,5 ,6 ,7 ,8 ] 这三方面.对POPs在水生生态系统环境行为及其影响因素的研究和分析相对较少.另外, 已有的有关POPs在水柱、沉积物、生物体和食物网中的研究相对独立, 缺少从生态系统的角度, 整体、全面、系统的探索 POPs 在不同相之间的环境行为、影响因素和互相联系.但实际上, POPs在水生生态系统中的环境行为是一个动态的整体过程, 在不同相之间的传递和分配是互相影响、互相联系的. ...
... ,4 ,5 ,6 ,7 ,8 ]这三方面.对POPs在水生生态系统环境行为及其影响因素的研究和分析相对较少.另外, 已有的有关POPs在水柱、沉积物、生物体和食物网中的研究相对独立, 缺少从生态系统的角度, 整体、全面、系统的探索 POPs 在不同相之间的环境行为、影响因素和互相联系.但实际上, POPs在水生生态系统中的环境行为是一个动态的整体过程, 在不同相之间的传递和分配是互相影响、互相联系的. ...
持久性有机污染物在渤海沉积物中的分布规律与来源解析[D]
4
2010
... 持久性有机污染物(POPs)具有持久性、生物蓄积性、高毒性、半挥发性和远距离传输的特点, 会对人类健康和生态安全产生严重危害.2001 年, 全球120多个国家签署了持久性有机污染物的斯德哥尔摩公约, 致力于削减 POPs 污染物.POPs 通过大气干湿沉降、地面径流和水-气界面直接交换进入水生生态系统, 使其成为 POPs在生态圈中传递的一个重要的“汇”.POPs 进入水体后, 在各种物理、化学、生物作用下发生迁移和转化.生物体通过鳃、皮肤等器官的被动扩散和捕食作用吸收 POPs, 在生物体内形成生物富集效应(biocon-centration), 并沿着食物链的传递造成生物放大(biomagnification), 使得鱼类等高等水生动物体内具有相当高浓度的 POPs, 并进一步通过渔业活动对人类健康造成严重威胁.因此, 研究 POPs在环境介质以及生物个体中的环境行为, 是揭示其在水生生态系统中归趋机制的基础, 对于准确地预测POPs 生物富集性的大小, 评价暴露风险, 研究POPs 的生物、生态毒性, 制定合理的环境基准, 实施科学的管控措施具有重要意义.鉴于此, 国内外对 POPs 在水、沉积物和土壤中的污染以及在不同环境介质和生物之间的迁移转化开展了大量的研 究[1 ,2 ,3 ,4 ,5 ,6 ,7 ,8 ] .目前国内有关水生生态系统中 POPs 的研究主要包括 POPs 在不同水生生态系统的水柱、沉积物、生物体内分布情况和浓度的研究[5 ,9 -15 ] , POPs在食物链中生物富集性的研究[1 ,2 ,3 ,4 ,5 ,6 ,7 ,8 ] , POPs的类型、性质、源解析和治理方法的研究[1 ,2 ,3 ,4 ,5 ,6 ,7 ,8 ] 这三方面.对POPs在水生生态系统环境行为及其影响因素的研究和分析相对较少.另外, 已有的有关POPs在水柱、沉积物、生物体和食物网中的研究相对独立, 缺少从生态系统的角度, 整体、全面、系统的探索 POPs 在不同相之间的环境行为、影响因素和互相联系.但实际上, POPs在水生生态系统中的环境行为是一个动态的整体过程, 在不同相之间的传递和分配是互相影响、互相联系的. ...
... [5 ,9 -15 ], POPs在食物链中生物富集性的研究[1 ,2 ,3 ,4 ,5 ,6 ,7 ,8 ] , POPs的类型、性质、源解析和治理方法的研究[1 ,2 ,3 ,4 ,5 ,6 ,7 ,8 ] 这三方面.对POPs在水生生态系统环境行为及其影响因素的研究和分析相对较少.另外, 已有的有关POPs在水柱、沉积物、生物体和食物网中的研究相对独立, 缺少从生态系统的角度, 整体、全面、系统的探索 POPs 在不同相之间的环境行为、影响因素和互相联系.但实际上, POPs在水生生态系统中的环境行为是一个动态的整体过程, 在不同相之间的传递和分配是互相影响、互相联系的. ...
... ,5 ,6 ,7 ,8 ], POPs的类型、性质、源解析和治理方法的研究[1 ,2 ,3 ,4 ,5 ,6 ,7 ,8 ] 这三方面.对POPs在水生生态系统环境行为及其影响因素的研究和分析相对较少.另外, 已有的有关POPs在水柱、沉积物、生物体和食物网中的研究相对独立, 缺少从生态系统的角度, 整体、全面、系统的探索 POPs 在不同相之间的环境行为、影响因素和互相联系.但实际上, POPs在水生生态系统中的环境行为是一个动态的整体过程, 在不同相之间的传递和分配是互相影响、互相联系的. ...
... ,5 ,6 ,7 ,8 ]这三方面.对POPs在水生生态系统环境行为及其影响因素的研究和分析相对较少.另外, 已有的有关POPs在水柱、沉积物、生物体和食物网中的研究相对独立, 缺少从生态系统的角度, 整体、全面、系统的探索 POPs 在不同相之间的环境行为、影响因素和互相联系.但实际上, POPs在水生生态系统中的环境行为是一个动态的整体过程, 在不同相之间的传递和分配是互相影响、互相联系的. ...
Residues, distributions, sources, and ecological risks of OCPs in the water from Lake Chaohu, China
3
2012
... 持久性有机污染物(POPs)具有持久性、生物蓄积性、高毒性、半挥发性和远距离传输的特点, 会对人类健康和生态安全产生严重危害.2001 年, 全球120多个国家签署了持久性有机污染物的斯德哥尔摩公约, 致力于削减 POPs 污染物.POPs 通过大气干湿沉降、地面径流和水-气界面直接交换进入水生生态系统, 使其成为 POPs在生态圈中传递的一个重要的“汇”.POPs 进入水体后, 在各种物理、化学、生物作用下发生迁移和转化.生物体通过鳃、皮肤等器官的被动扩散和捕食作用吸收 POPs, 在生物体内形成生物富集效应(biocon-centration), 并沿着食物链的传递造成生物放大(biomagnification), 使得鱼类等高等水生动物体内具有相当高浓度的 POPs, 并进一步通过渔业活动对人类健康造成严重威胁.因此, 研究 POPs在环境介质以及生物个体中的环境行为, 是揭示其在水生生态系统中归趋机制的基础, 对于准确地预测POPs 生物富集性的大小, 评价暴露风险, 研究POPs 的生物、生态毒性, 制定合理的环境基准, 实施科学的管控措施具有重要意义.鉴于此, 国内外对 POPs 在水、沉积物和土壤中的污染以及在不同环境介质和生物之间的迁移转化开展了大量的研 究[1 ,2 ,3 ,4 ,5 ,6 ,7 ,8 ] .目前国内有关水生生态系统中 POPs 的研究主要包括 POPs 在不同水生生态系统的水柱、沉积物、生物体内分布情况和浓度的研究[5 ,9 -15 ] , POPs在食物链中生物富集性的研究[1 ,2 ,3 ,4 ,5 ,6 ,7 ,8 ] , POPs的类型、性质、源解析和治理方法的研究[1 ,2 ,3 ,4 ,5 ,6 ,7 ,8 ] 这三方面.对POPs在水生生态系统环境行为及其影响因素的研究和分析相对较少.另外, 已有的有关POPs在水柱、沉积物、生物体和食物网中的研究相对独立, 缺少从生态系统的角度, 整体、全面、系统的探索 POPs 在不同相之间的环境行为、影响因素和互相联系.但实际上, POPs在水生生态系统中的环境行为是一个动态的整体过程, 在不同相之间的传递和分配是互相影响、互相联系的. ...
... ,6 ,7 ,8 ], POPs的类型、性质、源解析和治理方法的研究[1 ,2 ,3 ,4 ,5 ,6 ,7 ,8 ] 这三方面.对POPs在水生生态系统环境行为及其影响因素的研究和分析相对较少.另外, 已有的有关POPs在水柱、沉积物、生物体和食物网中的研究相对独立, 缺少从生态系统的角度, 整体、全面、系统的探索 POPs 在不同相之间的环境行为、影响因素和互相联系.但实际上, POPs在水生生态系统中的环境行为是一个动态的整体过程, 在不同相之间的传递和分配是互相影响、互相联系的. ...
... ,6 ,7 ,8 ]这三方面.对POPs在水生生态系统环境行为及其影响因素的研究和分析相对较少.另外, 已有的有关POPs在水柱、沉积物、生物体和食物网中的研究相对独立, 缺少从生态系统的角度, 整体、全面、系统的探索 POPs 在不同相之间的环境行为、影响因素和互相联系.但实际上, POPs在水生生态系统中的环境行为是一个动态的整体过程, 在不同相之间的传递和分配是互相影响、互相联系的. ...
Distributions, sources, and ecological risks of hexachlorocy-clohexanes in the sediments from Haihe Plain, Northern China
3
2013
... 持久性有机污染物(POPs)具有持久性、生物蓄积性、高毒性、半挥发性和远距离传输的特点, 会对人类健康和生态安全产生严重危害.2001 年, 全球120多个国家签署了持久性有机污染物的斯德哥尔摩公约, 致力于削减 POPs 污染物.POPs 通过大气干湿沉降、地面径流和水-气界面直接交换进入水生生态系统, 使其成为 POPs在生态圈中传递的一个重要的“汇”.POPs 进入水体后, 在各种物理、化学、生物作用下发生迁移和转化.生物体通过鳃、皮肤等器官的被动扩散和捕食作用吸收 POPs, 在生物体内形成生物富集效应(biocon-centration), 并沿着食物链的传递造成生物放大(biomagnification), 使得鱼类等高等水生动物体内具有相当高浓度的 POPs, 并进一步通过渔业活动对人类健康造成严重威胁.因此, 研究 POPs在环境介质以及生物个体中的环境行为, 是揭示其在水生生态系统中归趋机制的基础, 对于准确地预测POPs 生物富集性的大小, 评价暴露风险, 研究POPs 的生物、生态毒性, 制定合理的环境基准, 实施科学的管控措施具有重要意义.鉴于此, 国内外对 POPs 在水、沉积物和土壤中的污染以及在不同环境介质和生物之间的迁移转化开展了大量的研 究[1 ,2 ,3 ,4 ,5 ,6 ,7 ,8 ] .目前国内有关水生生态系统中 POPs 的研究主要包括 POPs 在不同水生生态系统的水柱、沉积物、生物体内分布情况和浓度的研究[5 ,9 -15 ] , POPs在食物链中生物富集性的研究[1 ,2 ,3 ,4 ,5 ,6 ,7 ,8 ] , POPs的类型、性质、源解析和治理方法的研究[1 ,2 ,3 ,4 ,5 ,6 ,7 ,8 ] 这三方面.对POPs在水生生态系统环境行为及其影响因素的研究和分析相对较少.另外, 已有的有关POPs在水柱、沉积物、生物体和食物网中的研究相对独立, 缺少从生态系统的角度, 整体、全面、系统的探索 POPs 在不同相之间的环境行为、影响因素和互相联系.但实际上, POPs在水生生态系统中的环境行为是一个动态的整体过程, 在不同相之间的传递和分配是互相影响、互相联系的. ...
... ,7 ,8 ], POPs的类型、性质、源解析和治理方法的研究[1 ,2 ,3 ,4 ,5 ,6 ,7 ,8 ] 这三方面.对POPs在水生生态系统环境行为及其影响因素的研究和分析相对较少.另外, 已有的有关POPs在水柱、沉积物、生物体和食物网中的研究相对独立, 缺少从生态系统的角度, 整体、全面、系统的探索 POPs 在不同相之间的环境行为、影响因素和互相联系.但实际上, POPs在水生生态系统中的环境行为是一个动态的整体过程, 在不同相之间的传递和分配是互相影响、互相联系的. ...
... ,7 ,8 ]这三方面.对POPs在水生生态系统环境行为及其影响因素的研究和分析相对较少.另外, 已有的有关POPs在水柱、沉积物、生物体和食物网中的研究相对独立, 缺少从生态系统的角度, 整体、全面、系统的探索 POPs 在不同相之间的环境行为、影响因素和互相联系.但实际上, POPs在水生生态系统中的环境行为是一个动态的整体过程, 在不同相之间的传递和分配是互相影响、互相联系的. ...
Measurements of persistent organic pollutants in Estonian ambient air (1990-2013)
3
2015
... 持久性有机污染物(POPs)具有持久性、生物蓄积性、高毒性、半挥发性和远距离传输的特点, 会对人类健康和生态安全产生严重危害.2001 年, 全球120多个国家签署了持久性有机污染物的斯德哥尔摩公约, 致力于削减 POPs 污染物.POPs 通过大气干湿沉降、地面径流和水-气界面直接交换进入水生生态系统, 使其成为 POPs在生态圈中传递的一个重要的“汇”.POPs 进入水体后, 在各种物理、化学、生物作用下发生迁移和转化.生物体通过鳃、皮肤等器官的被动扩散和捕食作用吸收 POPs, 在生物体内形成生物富集效应(biocon-centration), 并沿着食物链的传递造成生物放大(biomagnification), 使得鱼类等高等水生动物体内具有相当高浓度的 POPs, 并进一步通过渔业活动对人类健康造成严重威胁.因此, 研究 POPs在环境介质以及生物个体中的环境行为, 是揭示其在水生生态系统中归趋机制的基础, 对于准确地预测POPs 生物富集性的大小, 评价暴露风险, 研究POPs 的生物、生态毒性, 制定合理的环境基准, 实施科学的管控措施具有重要意义.鉴于此, 国内外对 POPs 在水、沉积物和土壤中的污染以及在不同环境介质和生物之间的迁移转化开展了大量的研 究[1 ,2 ,3 ,4 ,5 ,6 ,7 ,8 ] .目前国内有关水生生态系统中 POPs 的研究主要包括 POPs 在不同水生生态系统的水柱、沉积物、生物体内分布情况和浓度的研究[5 ,9 -15 ] , POPs在食物链中生物富集性的研究[1 ,2 ,3 ,4 ,5 ,6 ,7 ,8 ] , POPs的类型、性质、源解析和治理方法的研究[1 ,2 ,3 ,4 ,5 ,6 ,7 ,8 ] 这三方面.对POPs在水生生态系统环境行为及其影响因素的研究和分析相对较少.另外, 已有的有关POPs在水柱、沉积物、生物体和食物网中的研究相对独立, 缺少从生态系统的角度, 整体、全面、系统的探索 POPs 在不同相之间的环境行为、影响因素和互相联系.但实际上, POPs在水生生态系统中的环境行为是一个动态的整体过程, 在不同相之间的传递和分配是互相影响、互相联系的. ...
... ,8 ], POPs的类型、性质、源解析和治理方法的研究[1 ,2 ,3 ,4 ,5 ,6 ,7 ,8 ] 这三方面.对POPs在水生生态系统环境行为及其影响因素的研究和分析相对较少.另外, 已有的有关POPs在水柱、沉积物、生物体和食物网中的研究相对独立, 缺少从生态系统的角度, 整体、全面、系统的探索 POPs 在不同相之间的环境行为、影响因素和互相联系.但实际上, POPs在水生生态系统中的环境行为是一个动态的整体过程, 在不同相之间的传递和分配是互相影响、互相联系的. ...
... ,8 ]这三方面.对POPs在水生生态系统环境行为及其影响因素的研究和分析相对较少.另外, 已有的有关POPs在水柱、沉积物、生物体和食物网中的研究相对独立, 缺少从生态系统的角度, 整体、全面、系统的探索 POPs 在不同相之间的环境行为、影响因素和互相联系.但实际上, POPs在水生生态系统中的环境行为是一个动态的整体过程, 在不同相之间的传递和分配是互相影响、互相联系的. ...
湄洲湾水体中持久性有机污染物的污染特征与风险评估
2
2014
... 持久性有机污染物(POPs)具有持久性、生物蓄积性、高毒性、半挥发性和远距离传输的特点, 会对人类健康和生态安全产生严重危害.2001 年, 全球120多个国家签署了持久性有机污染物的斯德哥尔摩公约, 致力于削减 POPs 污染物.POPs 通过大气干湿沉降、地面径流和水-气界面直接交换进入水生生态系统, 使其成为 POPs在生态圈中传递的一个重要的“汇”.POPs 进入水体后, 在各种物理、化学、生物作用下发生迁移和转化.生物体通过鳃、皮肤等器官的被动扩散和捕食作用吸收 POPs, 在生物体内形成生物富集效应(biocon-centration), 并沿着食物链的传递造成生物放大(biomagnification), 使得鱼类等高等水生动物体内具有相当高浓度的 POPs, 并进一步通过渔业活动对人类健康造成严重威胁.因此, 研究 POPs在环境介质以及生物个体中的环境行为, 是揭示其在水生生态系统中归趋机制的基础, 对于准确地预测POPs 生物富集性的大小, 评价暴露风险, 研究POPs 的生物、生态毒性, 制定合理的环境基准, 实施科学的管控措施具有重要意义.鉴于此, 国内外对 POPs 在水、沉积物和土壤中的污染以及在不同环境介质和生物之间的迁移转化开展了大量的研 究[1 ,2 ,3 ,4 ,5 ,6 ,7 ,8 ] .目前国内有关水生生态系统中 POPs 的研究主要包括 POPs 在不同水生生态系统的水柱、沉积物、生物体内分布情况和浓度的研究[5 ,9 -15 ] , POPs在食物链中生物富集性的研究[1 ,2 ,3 ,4 ,5 ,6 ,7 ,8 ] , POPs的类型、性质、源解析和治理方法的研究[1 ,2 ,3 ,4 ,5 ,6 ,7 ,8 ] 这三方面.对POPs在水生生态系统环境行为及其影响因素的研究和分析相对较少.另外, 已有的有关POPs在水柱、沉积物、生物体和食物网中的研究相对独立, 缺少从生态系统的角度, 整体、全面、系统的探索 POPs 在不同相之间的环境行为、影响因素和互相联系.但实际上, POPs在水生生态系统中的环境行为是一个动态的整体过程, 在不同相之间的传递和分配是互相影响、互相联系的. ...
... POPs 进入水体后, 会在各种物理、化学、生物因素影响下, 在水柱和沉积物中发生迁移和转化(图 1 ).水柱和沉积物是水生生物体生活和栖息场所, 也是其食物和能量来源.生物体通过被动扩散和摄食等活动从环境介质中吸收并富集 POPs, 并进一步沿食物网传递.因此, 研究 POPs 在水柱和沉积物等介质中的环境行为, 是研究 POPs 在水生生态系统中传递的基础, 是决定 POPs 在水生生物和食物网中生物富集和生物放大的前提.POPs 通过地表径流、大气干湿沉降、大气-水界面扩散等作用进入水体[9 ] , 其环境行为和过程受到诸多因素的影响, 如温度、风速、悬浮颗粒物、浮游植物、腐殖质含量、沉积物再悬浮和微生物等[2 ,23 -24 ] . ...
持久性有机污染物在东海泥质区的分布与沉积记录[D]
2005
渤海湾西岸典型持久性有机污染物的环境行为初步研究[D]
2010
持久性有机污染物在贡湖沉积物和水体中的分布特征及源解析[D]
2010
Butyltin compounds distribution in the coastal waters of Bohai Bay, People’s Republic of China. Bulletin of Environm-ental Contamination and
2004
Fate of DDT-related compounds in Bohai Bay and its adjacent Haihe Basin, north China
2005
影响沉积物-水界面持久性有机污染物迁移行为的因素研究
1
2011
... 持久性有机污染物(POPs)具有持久性、生物蓄积性、高毒性、半挥发性和远距离传输的特点, 会对人类健康和生态安全产生严重危害.2001 年, 全球120多个国家签署了持久性有机污染物的斯德哥尔摩公约, 致力于削减 POPs 污染物.POPs 通过大气干湿沉降、地面径流和水-气界面直接交换进入水生生态系统, 使其成为 POPs在生态圈中传递的一个重要的“汇”.POPs 进入水体后, 在各种物理、化学、生物作用下发生迁移和转化.生物体通过鳃、皮肤等器官的被动扩散和捕食作用吸收 POPs, 在生物体内形成生物富集效应(biocon-centration), 并沿着食物链的传递造成生物放大(biomagnification), 使得鱼类等高等水生动物体内具有相当高浓度的 POPs, 并进一步通过渔业活动对人类健康造成严重威胁.因此, 研究 POPs在环境介质以及生物个体中的环境行为, 是揭示其在水生生态系统中归趋机制的基础, 对于准确地预测POPs 生物富集性的大小, 评价暴露风险, 研究POPs 的生物、生态毒性, 制定合理的环境基准, 实施科学的管控措施具有重要意义.鉴于此, 国内外对 POPs 在水、沉积物和土壤中的污染以及在不同环境介质和生物之间的迁移转化开展了大量的研 究[1 ,2 ,3 ,4 ,5 ,6 ,7 ,8 ] .目前国内有关水生生态系统中 POPs 的研究主要包括 POPs 在不同水生生态系统的水柱、沉积物、生物体内分布情况和浓度的研究[5 ,9 -15 ] , POPs在食物链中生物富集性的研究[1 ,2 ,3 ,4 ,5 ,6 ,7 ,8 ] , POPs的类型、性质、源解析和治理方法的研究[1 ,2 ,3 ,4 ,5 ,6 ,7 ,8 ] 这三方面.对POPs在水生生态系统环境行为及其影响因素的研究和分析相对较少.另外, 已有的有关POPs在水柱、沉积物、生物体和食物网中的研究相对独立, 缺少从生态系统的角度, 整体、全面、系统的探索 POPs 在不同相之间的环境行为、影响因素和互相联系.但实际上, POPs在水生生态系统中的环境行为是一个动态的整体过程, 在不同相之间的传递和分配是互相影响、互相联系的. ...
两类持久性有机污染物和重金属在南四湖食物链中的分布和生物积累[D]
2014
Characterization of trophic transfer for PCDD/Fs and coplanar PCBs in the marine food web of Bohai Bay, North China
2005
J, Wan Y, et al. Trophic magnification of triphenyltin in marine food web: comparison to tributyltin
2006
持久性有机污染物在食物链中积累与放大研究进展
2008
持久性有机污染物的水污染控制: 吸附富集、生物降解与过程分析
2011
持久性有机污染物(POPs)生物修复研究进展
2006
中国淡水环境中典型持久性有机污染物(POPs)的污染现状与分布特征
2013
Lake productivity and water chemistry as governors of the uptake of POPs in fish
6
1992
... POPs 进入水体后, 会在各种物理、化学、生物因素影响下, 在水柱和沉积物中发生迁移和转化(图 1 ).水柱和沉积物是水生生物体生活和栖息场所, 也是其食物和能量来源.生物体通过被动扩散和摄食等活动从环境介质中吸收并富集 POPs, 并进一步沿食物网传递.因此, 研究 POPs 在水柱和沉积物等介质中的环境行为, 是研究 POPs 在水生生态系统中传递的基础, 是决定 POPs 在水生生物和食物网中生物富集和生物放大的前提.POPs 通过地表径流、大气干湿沉降、大气-水界面扩散等作用进入水体[9 ] , 其环境行为和过程受到诸多因素的影响, 如温度、风速、悬浮颗粒物、浮游植物、腐殖质含量、沉积物再悬浮和微生物等[2 ,23 -24 ] . ...
... POPs 在水-气界面的扩散交换会受到风速和温度等因素的影响.风速通过改变大气-水表面之间的质量传质系数影响 POPs 的扩散作用; 风速越高, 温度越高, 水面 POPs 的挥发和扩散加快.水体的生产力、浮游植物和微生物的吸附以及颗粒物的沉降等也能够影响水-气界面的 POPs 转移, 进而影响POPs 在水生生态系统中的归趋.研究表明, 当水体发生富营养化时, 浮游植物的生长率和生物量会大大增加, 浮游植物的吸收会导致水柱中 PCBs 浓度降低, 打破 PCBs 在水-气界面的平衡状态, 促进PCBs从大气向水柱转移[2 ] .当浮游植物的生物量翻倍时, 大气-水柱之间某些有机污染物的交换通量能够增加 32%~37%, 一般低氯代污染物的增加量要比高氯代污染物的增加量多.在水柱中, POPs被颗粒物(主要是浮游植物)大量吸附和吸收后, 随着颗粒物的沉降向底栖生境转移, 增加了沉积物中POPs的浓度[24 ] .因此, 沉积物成为POPs进入水环境以后的主要蓄积场所和归宿, 沉积物中的 POPs浓度通常要比相应水柱中的浓度高几个, 甚至十几个数量级[25 ] .此外, 腐殖质的含量和成分也是影响水柱中及生物体内 POPs 浓度的重要因素.Carter等[26 ] 发现, 腐殖质吸附 POPs 的程度受到腐殖质来源、类型、pH 值、Ca 浓度、离子化程度和腐殖质浓度大小的影响.进一步的研究表明, 随着水体中腐殖质增加, POPs 浓度会不断下降, 原因是腐殖质能够大量吸附 POPs, 降低水体以及生物体内 POPs的浓度, 且对于疏水性较强的 POPs 的影响相对更大[23 ] . ...
... Physiological features that influence the bioaccumulation
生理特征 对BAFs的影响 主要文献 生长率 生长率加快, 导致“生长稀释”, 降低水柱、浮游生物等生物体内的POPs浓度 [23 ] 性别 雌性生物通过产卵, 降低体内POPs浓度 [23 , 43 -44 ] 脂质 脂质与体内POPs浓度呈正相关 [33 , 45 ] 生物转化 生物转化会降低POPs浓度, 不同生物降解不同种类POPs的能力不同 [35 , 38 -39 ] 体型大小 有研究认为, 体型增大, 由于吸收、储存、转化等因素, 生物体对POPs的生物富集性降低; [46 ,47 ,48 ,49 ,50 ,51 ]
生长率的提高会降低生物体内POPs 浓度.在夏季富营养化的湖泊中, 浮游植物生长最快, 生物量增加, 此时体内 POPs 浓度较低, 尤其是高氯代PCBs 等疏水性较强的化合物.原因在于: 1) 浮游植物大量生长、生物量增大, 形成稀释效应; 2) 浮游植物对 POPs 的大量吸收, 打破了水-气界面的平衡分配, 降低了水柱中溶解态POPs的浓度; 3) 浮游植物对 POPs 的吸收分为两个主要的步骤, 首先是POPs 被吸附在浮游植物的表面(较快), 之后 POPs从细胞的表面转移到细胞内部(较慢), 分子越大, 脂溶性越强, 转移越慢.当浮游植物生长率较快时, POPs 在浮游植物和水柱之间分配失衡.因此浮游植物的生长率越快, 生物量越大, POPs 的脂溶性越强, 生长稀释效应就越大.不仅浮游植物如此, 鱼类等其他生物当生长率大大提高的时候, 其体内的POPs浓度也会下降[23 ] . ...
... [
23 ,
43 -
44 ]
脂质 脂质与体内POPs浓度呈正相关 [33 , 45 ] 生物转化 生物转化会降低POPs浓度, 不同生物降解不同种类POPs的能力不同 [35 , 38 -39 ] 体型大小 有研究认为, 体型增大, 由于吸收、储存、转化等因素, 生物体对POPs的生物富集性降低; [46 ,47 ,48 ,49 ,50 ,51 ] 生长率的提高会降低生物体内POPs 浓度.在夏季富营养化的湖泊中, 浮游植物生长最快, 生物量增加, 此时体内 POPs 浓度较低, 尤其是高氯代PCBs 等疏水性较强的化合物.原因在于: 1) 浮游植物大量生长、生物量增大, 形成稀释效应; 2) 浮游植物对 POPs 的大量吸收, 打破了水-气界面的平衡分配, 降低了水柱中溶解态POPs的浓度; 3) 浮游植物对 POPs 的吸收分为两个主要的步骤, 首先是POPs 被吸附在浮游植物的表面(较快), 之后 POPs从细胞的表面转移到细胞内部(较慢), 分子越大, 脂溶性越强, 转移越慢.当浮游植物生长率较快时, POPs 在浮游植物和水柱之间分配失衡.因此浮游植物的生长率越快, 生物量越大, POPs 的脂溶性越强, 生长稀释效应就越大.不仅浮游植物如此, 鱼类等其他生物当生长率大大提高的时候, 其体内的POPs浓度也会下降[23 ] . ...
... 生长率的提高会降低生物体内POPs 浓度.在夏季富营养化的湖泊中, 浮游植物生长最快, 生物量增加, 此时体内 POPs 浓度较低, 尤其是高氯代PCBs 等疏水性较强的化合物.原因在于: 1) 浮游植物大量生长、生物量增大, 形成稀释效应; 2) 浮游植物对 POPs 的大量吸收, 打破了水-气界面的平衡分配, 降低了水柱中溶解态POPs的浓度; 3) 浮游植物对 POPs 的吸收分为两个主要的步骤, 首先是POPs 被吸附在浮游植物的表面(较快), 之后 POPs从细胞的表面转移到细胞内部(较慢), 分子越大, 脂溶性越强, 转移越慢.当浮游植物生长率较快时, POPs 在浮游植物和水柱之间分配失衡.因此浮游植物的生长率越快, 生物量越大, POPs 的脂溶性越强, 生长稀释效应就越大.不仅浮游植物如此, 鱼类等其他生物当生长率大大提高的时候, 其体内的POPs浓度也会下降[23 ] . ...
... 雄性生物体内的 POPs 浓度高于雌性, 这是因为雌性生物会通过产卵降低体内 POPs 的浓度.研究发现, 雌性梭子鱼体内的 POPs 浓度随着年龄、体重和体型的增大而线性下降, 这是由雌鱼的季节性产卵造成的, 卵中含有的脂质含量比肌肉高10 倍, 其污染物的浓度也相应地高 10 倍左右, 而雄性通过性器官排泄的污染物较少[23 ] .此外, McManus 等[43 ] 发现, 雌性桡足类动物在产卵过程中, 体内的 PCBs 浓度也下降较快, 同样是由于很大一部分 PCBs 转移到富含脂肪的卵上.Lotufo[44 ] 将小型的底栖桡足类暴露在荧蒽中, 发现有 50%的化合物留在卵中, 通过产卵排出体外. ...
Coupling of phytoplankton uptake and air-water exchange of persistent organic pollutants
2
1999
... POPs 进入水体后, 会在各种物理、化学、生物因素影响下, 在水柱和沉积物中发生迁移和转化(图 1 ).水柱和沉积物是水生生物体生活和栖息场所, 也是其食物和能量来源.生物体通过被动扩散和摄食等活动从环境介质中吸收并富集 POPs, 并进一步沿食物网传递.因此, 研究 POPs 在水柱和沉积物等介质中的环境行为, 是研究 POPs 在水生生态系统中传递的基础, 是决定 POPs 在水生生物和食物网中生物富集和生物放大的前提.POPs 通过地表径流、大气干湿沉降、大气-水界面扩散等作用进入水体[9 ] , 其环境行为和过程受到诸多因素的影响, 如温度、风速、悬浮颗粒物、浮游植物、腐殖质含量、沉积物再悬浮和微生物等[2 ,23 -24 ] . ...
... POPs 在水-气界面的扩散交换会受到风速和温度等因素的影响.风速通过改变大气-水表面之间的质量传质系数影响 POPs 的扩散作用; 风速越高, 温度越高, 水面 POPs 的挥发和扩散加快.水体的生产力、浮游植物和微生物的吸附以及颗粒物的沉降等也能够影响水-气界面的 POPs 转移, 进而影响POPs 在水生生态系统中的归趋.研究表明, 当水体发生富营养化时, 浮游植物的生长率和生物量会大大增加, 浮游植物的吸收会导致水柱中 PCBs 浓度降低, 打破 PCBs 在水-气界面的平衡状态, 促进PCBs从大气向水柱转移[2 ] .当浮游植物的生物量翻倍时, 大气-水柱之间某些有机污染物的交换通量能够增加 32%~37%, 一般低氯代污染物的增加量要比高氯代污染物的增加量多.在水柱中, POPs被颗粒物(主要是浮游植物)大量吸附和吸收后, 随着颗粒物的沉降向底栖生境转移, 增加了沉积物中POPs的浓度[24 ] .因此, 沉积物成为POPs进入水环境以后的主要蓄积场所和归宿, 沉积物中的 POPs浓度通常要比相应水柱中的浓度高几个, 甚至十几个数量级[25 ] .此外, 腐殖质的含量和成分也是影响水柱中及生物体内 POPs 浓度的重要因素.Carter等[26 ] 发现, 腐殖质吸附 POPs 的程度受到腐殖质来源、类型、pH 值、Ca 浓度、离子化程度和腐殖质浓度大小的影响.进一步的研究表明, 随着水体中腐殖质增加, POPs 浓度会不断下降, 原因是腐殖质能够大量吸附 POPs, 降低水体以及生物体内 POPs的浓度, 且对于疏水性较强的 POPs 的影响相对更大[23 ] . ...
有机氯农药在湖泊水体和沉积物中的污染特征及动力学特征: 以洪湖为例[D]
2
2007
... POPs 在水-气界面的扩散交换会受到风速和温度等因素的影响.风速通过改变大气-水表面之间的质量传质系数影响 POPs 的扩散作用; 风速越高, 温度越高, 水面 POPs 的挥发和扩散加快.水体的生产力、浮游植物和微生物的吸附以及颗粒物的沉降等也能够影响水-气界面的 POPs 转移, 进而影响POPs 在水生生态系统中的归趋.研究表明, 当水体发生富营养化时, 浮游植物的生长率和生物量会大大增加, 浮游植物的吸收会导致水柱中 PCBs 浓度降低, 打破 PCBs 在水-气界面的平衡状态, 促进PCBs从大气向水柱转移[2 ] .当浮游植物的生物量翻倍时, 大气-水柱之间某些有机污染物的交换通量能够增加 32%~37%, 一般低氯代污染物的增加量要比高氯代污染物的增加量多.在水柱中, POPs被颗粒物(主要是浮游植物)大量吸附和吸收后, 随着颗粒物的沉降向底栖生境转移, 增加了沉积物中POPs的浓度[24 ] .因此, 沉积物成为POPs进入水环境以后的主要蓄积场所和归宿, 沉积物中的 POPs浓度通常要比相应水柱中的浓度高几个, 甚至十几个数量级[25 ] .此外, 腐殖质的含量和成分也是影响水柱中及生物体内 POPs 浓度的重要因素.Carter等[26 ] 发现, 腐殖质吸附 POPs 的程度受到腐殖质来源、类型、pH 值、Ca 浓度、离子化程度和腐殖质浓度大小的影响.进一步的研究表明, 随着水体中腐殖质增加, POPs 浓度会不断下降, 原因是腐殖质能够大量吸附 POPs, 降低水体以及生物体内 POPs的浓度, 且对于疏水性较强的 POPs 的影响相对更大[23 ] . ...
... 沉积物中的 POPs 在一定条件下会发生各种转化, 或从颗粒物上解吸, 再次进入水柱中.微生物分解是影响 POPs 含量和分布类型的重要因素, 且在不同的环境条件下, 微生物对 POPs 的降解活性和转化产物不同.例如, 对鸭川河 3 个不同站点沉积物中的林丹和六氯苯的降解研究发现, 微生物对3个站点的六氯苯(HCBs)几乎是同时开始降解的, 但对于反-林丹(TC)和顺-林丹(CC), 不同采样点开始降解的时间有很大差异.当实验结束时, 不同种类 POPs 的降解率差别很大, 分别是 HCBs>TC> CC.同时, 微生物对POPs的降解也受到环境理化条件的影响.例如, 沉积物中的有机碳(TOC)含量越高、有机污染越严重, 微生物对 POPs 的降解速率越高[31 ] .DDT 在厌氧条件下被微生物分解为DDD, 而在好氧条件下转为 DDE[32 ] .透光层的颗粒物在沉降过程中, 有机质不断降解, 浓度下降, 但 POPs 的浓度却不断升高, 在颗粒物沉降到沉积物表面之后, 大部分有机质被微生物降解, 此时大部分POPs 被释放出来, 再次进入水柱中循环.除微生物外, 温度等物理条件同样也是影响沉积物中POPs 再循环的重要因素.研究发现, 随着温度的升高, 沉积物中有机氯农药的释放明显加强, 且在紊动条件下的释放速率和强度远远大于静止条件 下[25 ] .沉积物中的POPs 除通过解吸释放到水柱中外, 还可以通过沉积物的再悬浮进入水柱中.湖泊在秋季分层结束时, 沉积物中的 POPs 会随着颗粒物的再悬浮再次进入水柱中.研究证实, 现场调查到的水柱中POPs沉降通量会比大气沉降通量高1~2 个数量级, 就是由沉积物的再悬浮造成的[33 ] . ...
1
1982
... POPs 在水-气界面的扩散交换会受到风速和温度等因素的影响.风速通过改变大气-水表面之间的质量传质系数影响 POPs 的扩散作用; 风速越高, 温度越高, 水面 POPs 的挥发和扩散加快.水体的生产力、浮游植物和微生物的吸附以及颗粒物的沉降等也能够影响水-气界面的 POPs 转移, 进而影响POPs 在水生生态系统中的归趋.研究表明, 当水体发生富营养化时, 浮游植物的生长率和生物量会大大增加, 浮游植物的吸收会导致水柱中 PCBs 浓度降低, 打破 PCBs 在水-气界面的平衡状态, 促进PCBs从大气向水柱转移[2 ] .当浮游植物的生物量翻倍时, 大气-水柱之间某些有机污染物的交换通量能够增加 32%~37%, 一般低氯代污染物的增加量要比高氯代污染物的增加量多.在水柱中, POPs被颗粒物(主要是浮游植物)大量吸附和吸收后, 随着颗粒物的沉降向底栖生境转移, 增加了沉积物中POPs的浓度[24 ] .因此, 沉积物成为POPs进入水环境以后的主要蓄积场所和归宿, 沉积物中的 POPs浓度通常要比相应水柱中的浓度高几个, 甚至十几个数量级[25 ] .此外, 腐殖质的含量和成分也是影响水柱中及生物体内 POPs 浓度的重要因素.Carter等[26 ] 发现, 腐殖质吸附 POPs 的程度受到腐殖质来源、类型、pH 值、Ca 浓度、离子化程度和腐殖质浓度大小的影响.进一步的研究表明, 随着水体中腐殖质增加, POPs 浓度会不断下降, 原因是腐殖质能够大量吸附 POPs, 降低水体以及生物体内 POPs的浓度, 且对于疏水性较强的 POPs 的影响相对更大[23 ] . ...
Associations of chlorinated hydrocarbons with fine Particles and humic substan-ces innearshore surficial sediments
1
1976
... 由于沉积物的化学成分和结构形态不同, 它们与 POPs 相互作用决定了 POPs 的稳定程度、迁移速率和生物有效性的差异很大.砂质沉积物中的POPs 浓度较小, 而泥质沉积物中由于有机质含量高, 吸附能力强, POPs 的浓度较大.沉积物中有机质的组成类型也对 POPs 的吸附有很大的影响, 粒径越小的颗粒物, 其有机碳含量越高, 吸附 POPs的能力越强[27 ,28 ,29 ] .研究发现, 沉积物中总有机碳(TOC)含量与 PAHs 含量显著相关, 沉积物的粒径与OCPs和 PCBs 的含量之间有很好的相关性[30 ] . ...
Sorption of hydrophobic Pollutants on nature sediments
1
1979
... 由于沉积物的化学成分和结构形态不同, 它们与 POPs 相互作用决定了 POPs 的稳定程度、迁移速率和生物有效性的差异很大.砂质沉积物中的POPs 浓度较小, 而泥质沉积物中由于有机质含量高, 吸附能力强, POPs 的浓度较大.沉积物中有机质的组成类型也对 POPs 的吸附有很大的影响, 粒径越小的颗粒物, 其有机碳含量越高, 吸附 POPs的能力越强[27 ,28 ,29 ] .研究发现, 沉积物中总有机碳(TOC)含量与 PAHs 含量显著相关, 沉积物的粒径与OCPs和 PCBs 的含量之间有很好的相关性[30 ] . ...
Water dispersible soil organic mineral particles: I. Carbon and nitrogen distribution
1
1979
... 由于沉积物的化学成分和结构形态不同, 它们与 POPs 相互作用决定了 POPs 的稳定程度、迁移速率和生物有效性的差异很大.砂质沉积物中的POPs 浓度较小, 而泥质沉积物中由于有机质含量高, 吸附能力强, POPs 的浓度较大.沉积物中有机质的组成类型也对 POPs 的吸附有很大的影响, 粒径越小的颗粒物, 其有机碳含量越高, 吸附 POPs的能力越强[27 ,28 ,29 ] .研究发现, 沉积物中总有机碳(TOC)含量与 PAHs 含量显著相关, 沉积物的粒径与OCPs和 PCBs 的含量之间有很好的相关性[30 ] . ...
1
2011
... 由于沉积物的化学成分和结构形态不同, 它们与 POPs 相互作用决定了 POPs 的稳定程度、迁移速率和生物有效性的差异很大.砂质沉积物中的POPs 浓度较小, 而泥质沉积物中由于有机质含量高, 吸附能力强, POPs 的浓度较大.沉积物中有机质的组成类型也对 POPs 的吸附有很大的影响, 粒径越小的颗粒物, 其有机碳含量越高, 吸附 POPs的能力越强[27 ,28 ,29 ] .研究发现, 沉积物中总有机碳(TOC)含量与 PAHs 含量显著相关, 沉积物的粒径与OCPs和 PCBs 的含量之间有很好的相关性[30 ] . ...
Biodegradation of chlordane and hexachorobenzenes in river sedimen
1
2007
... 沉积物中的 POPs 在一定条件下会发生各种转化, 或从颗粒物上解吸, 再次进入水柱中.微生物分解是影响 POPs 含量和分布类型的重要因素, 且在不同的环境条件下, 微生物对 POPs 的降解活性和转化产物不同.例如, 对鸭川河 3 个不同站点沉积物中的林丹和六氯苯的降解研究发现, 微生物对3个站点的六氯苯(HCBs)几乎是同时开始降解的, 但对于反-林丹(TC)和顺-林丹(CC), 不同采样点开始降解的时间有很大差异.当实验结束时, 不同种类 POPs 的降解率差别很大, 分别是 HCBs>TC> CC.同时, 微生物对POPs的降解也受到环境理化条件的影响.例如, 沉积物中的有机碳(TOC)含量越高、有机污染越严重, 微生物对 POPs 的降解速率越高[31 ] .DDT 在厌氧条件下被微生物分解为DDD, 而在好氧条件下转为 DDE[32 ] .透光层的颗粒物在沉降过程中, 有机质不断降解, 浓度下降, 但 POPs 的浓度却不断升高, 在颗粒物沉降到沉积物表面之后, 大部分有机质被微生物降解, 此时大部分POPs 被释放出来, 再次进入水柱中循环.除微生物外, 温度等物理条件同样也是影响沉积物中POPs 再循环的重要因素.研究发现, 随着温度的升高, 沉积物中有机氯农药的释放明显加强, 且在紊动条件下的释放速率和强度远远大于静止条件 下[25 ] .沉积物中的POPs 除通过解吸释放到水柱中外, 还可以通过沉积物的再悬浮进入水柱中.湖泊在秋季分层结束时, 沉积物中的 POPs 会随着颗粒物的再悬浮再次进入水柱中.研究证实, 现场调查到的水柱中POPs沉降通量会比大气沉降通量高1~2 个数量级, 就是由沉积物的再悬浮造成的[33 ] . ...
Unusual Persistence of DDT in some Western USA soils
1
1992
... 沉积物中的 POPs 在一定条件下会发生各种转化, 或从颗粒物上解吸, 再次进入水柱中.微生物分解是影响 POPs 含量和分布类型的重要因素, 且在不同的环境条件下, 微生物对 POPs 的降解活性和转化产物不同.例如, 对鸭川河 3 个不同站点沉积物中的林丹和六氯苯的降解研究发现, 微生物对3个站点的六氯苯(HCBs)几乎是同时开始降解的, 但对于反-林丹(TC)和顺-林丹(CC), 不同采样点开始降解的时间有很大差异.当实验结束时, 不同种类 POPs 的降解率差别很大, 分别是 HCBs>TC> CC.同时, 微生物对POPs的降解也受到环境理化条件的影响.例如, 沉积物中的有机碳(TOC)含量越高、有机污染越严重, 微生物对 POPs 的降解速率越高[31 ] .DDT 在厌氧条件下被微生物分解为DDD, 而在好氧条件下转为 DDE[32 ] .透光层的颗粒物在沉降过程中, 有机质不断降解, 浓度下降, 但 POPs 的浓度却不断升高, 在颗粒物沉降到沉积物表面之后, 大部分有机质被微生物降解, 此时大部分POPs 被释放出来, 再次进入水柱中循环.除微生物外, 温度等物理条件同样也是影响沉积物中POPs 再循环的重要因素.研究发现, 随着温度的升高, 沉积物中有机氯农药的释放明显加强, 且在紊动条件下的释放速率和强度远远大于静止条件 下[25 ] .沉积物中的POPs 除通过解吸释放到水柱中外, 还可以通过沉积物的再悬浮进入水柱中.湖泊在秋季分层结束时, 沉积物中的 POPs 会随着颗粒物的再悬浮再次进入水柱中.研究证实, 现场调查到的水柱中POPs沉降通量会比大气沉降通量高1~2 个数量级, 就是由沉积物的再悬浮造成的[33 ] . ...
Persistent organic pollutants (POPs) in pelagic system
3
2000
... 沉积物中的 POPs 在一定条件下会发生各种转化, 或从颗粒物上解吸, 再次进入水柱中.微生物分解是影响 POPs 含量和分布类型的重要因素, 且在不同的环境条件下, 微生物对 POPs 的降解活性和转化产物不同.例如, 对鸭川河 3 个不同站点沉积物中的林丹和六氯苯的降解研究发现, 微生物对3个站点的六氯苯(HCBs)几乎是同时开始降解的, 但对于反-林丹(TC)和顺-林丹(CC), 不同采样点开始降解的时间有很大差异.当实验结束时, 不同种类 POPs 的降解率差别很大, 分别是 HCBs>TC> CC.同时, 微生物对POPs的降解也受到环境理化条件的影响.例如, 沉积物中的有机碳(TOC)含量越高、有机污染越严重, 微生物对 POPs 的降解速率越高[31 ] .DDT 在厌氧条件下被微生物分解为DDD, 而在好氧条件下转为 DDE[32 ] .透光层的颗粒物在沉降过程中, 有机质不断降解, 浓度下降, 但 POPs 的浓度却不断升高, 在颗粒物沉降到沉积物表面之后, 大部分有机质被微生物降解, 此时大部分POPs 被释放出来, 再次进入水柱中循环.除微生物外, 温度等物理条件同样也是影响沉积物中POPs 再循环的重要因素.研究发现, 随着温度的升高, 沉积物中有机氯农药的释放明显加强, 且在紊动条件下的释放速率和强度远远大于静止条件 下[25 ] .沉积物中的POPs 除通过解吸释放到水柱中外, 还可以通过沉积物的再悬浮进入水柱中.湖泊在秋季分层结束时, 沉积物中的 POPs 会随着颗粒物的再悬浮再次进入水柱中.研究证实, 现场调查到的水柱中POPs沉降通量会比大气沉降通量高1~2 个数量级, 就是由沉积物的再悬浮造成的[33 ] . ...
... Physiological features that influence the bioaccumulation
生理特征 对BAFs的影响 主要文献 生长率 生长率加快, 导致“生长稀释”, 降低水柱、浮游生物等生物体内的POPs浓度 [23 ] 性别 雌性生物通过产卵, 降低体内POPs浓度 [23 , 43 -44 ] 脂质 脂质与体内POPs浓度呈正相关 [33 , 45 ] 生物转化 生物转化会降低POPs浓度, 不同生物降解不同种类POPs的能力不同 [35 , 38 -39 ] 体型大小 有研究认为, 体型增大, 由于吸收、储存、转化等因素, 生物体对POPs的生物富集性降低; [46 ,47 ,48 ,49 ,50 ,51 ]
生长率的提高会降低生物体内POPs 浓度.在夏季富营养化的湖泊中, 浮游植物生长最快, 生物量增加, 此时体内 POPs 浓度较低, 尤其是高氯代PCBs 等疏水性较强的化合物.原因在于: 1) 浮游植物大量生长、生物量增大, 形成稀释效应; 2) 浮游植物对 POPs 的大量吸收, 打破了水-气界面的平衡分配, 降低了水柱中溶解态POPs的浓度; 3) 浮游植物对 POPs 的吸收分为两个主要的步骤, 首先是POPs 被吸附在浮游植物的表面(较快), 之后 POPs从细胞的表面转移到细胞内部(较慢), 分子越大, 脂溶性越强, 转移越慢.当浮游植物生长率较快时, POPs 在浮游植物和水柱之间分配失衡.因此浮游植物的生长率越快, 生物量越大, POPs 的脂溶性越强, 生长稀释效应就越大.不仅浮游植物如此, 鱼类等其他生物当生长率大大提高的时候, 其体内的POPs浓度也会下降[23 ] . ...
... 生物体的脂质含量与其体内的 POPs浓度具有显著的正相关关系.Kidd 等[45 ] 发现 DDT 和 DDE的浓度与生物体脂质含量显著正相关, 且浮游生物要比相同营养级的底栖生物体内的 DDT 浓度高, 推测原因是前者的脂质含量比后者高.另外, 在寡营养湖泊中, 浮游生物、鱼类等生物体内的 POPs浓度比富营养条件下高很多, 除上面提到的生物稀释等因素以外, 寡营养条件下生物体内脂质含量更高也是一个重要原因[33 ] . ...
Critical review and recommended values for the physical-chemical pro-perty data of 15 Polycyclic Aromatic Hydrocarbons at 25°C
1
2010
... POPs 的理化性质主要包括种类、分子大小、分子结构、亲脂性、卤代程度等, 但它们之间互相关联.POPs 的苯环数量越多, 卤代程度越高, 分子越大, 亲脂性越强.在不同情况下, 这几方面因素在 POPs 生物富集过程起的作用不同, 有时是协同作用, 有时作用是相反的[3 ,34 -35 ] .例如, 氯代程度越高, 分子越大, 亲脂性越强, 因此生物富集性会越高; 但氯代程度越大, 分子也会变大, 由于阻碍作用又会降低生物富集性.因此, 在研究 POPs 的生物富集性时, 尤其是同一类 POPs 的各种同系物的生物富集性时, 需同时评价这几类因素的影响(图2 ). ...
Trophodyna- mic behavior of hydrophobic organic contaminants in the aquatic food web of a tidal river
6
2014
... POPs 的理化性质主要包括种类、分子大小、分子结构、亲脂性、卤代程度等, 但它们之间互相关联.POPs 的苯环数量越多, 卤代程度越高, 分子越大, 亲脂性越强.在不同情况下, 这几方面因素在 POPs 生物富集过程起的作用不同, 有时是协同作用, 有时作用是相反的[3 ,34 -35 ] .例如, 氯代程度越高, 分子越大, 亲脂性越强, 因此生物富集性会越高; 但氯代程度越大, 分子也会变大, 由于阻碍作用又会降低生物富集性.因此, 在研究 POPs 的生物富集性时, 尤其是同一类 POPs 的各种同系物的生物富集性时, 需同时评价这几类因素的影响(图2 ). ...
... 2) POPs 的种类、亲脂性和结构会影响其生物集性及其在生物体内的生物转化和代谢分解.Mizukawa 等[3 ] 分析日本东京湾中双壳类、螃蟹、鱼、海水以及沉积物中所含的 20 种 PBDEs 和PCBs 的生物富集性, 发现 PBDEs 和 PCBs 的生物浓缩因子(BCFs)随着 K ow 的增加而增加, 但是当lgK ow >7以后, BCFs 随着 K ow 的增加而下降.PCBs和大多数的 PBDEs 同系物存在生物放大作用, 但另外几种 PBDEs 同系物不存在生物放大作用, 其原因可能是在生物体内被生物转化或代谢分解了.Hoekstra等[37 ] 研究了几类有机污染物在阿拉斯加和加拿大极地地区的桡足类生物体内的生物富集性, 其大小顺序为: 毒杀芬>PCBs>HCHs>DDT> CHLOR >CIBz.毒杀芬和 PCBs 生物富集性之所以较大, 是因为生物体内细胞色素 P-4502B 同工酶对这两类物质的生物转化量较少.另有研究发现, 疏水性相同的PAHs 的生物富集性小于 PCBs, 因为PAHs 在甲壳类和其他无脊椎动物体内较容易被代谢而排出[35 ,38 ] . ...
... 3) 分子结构和卤代程度会影响 POPs 的生物富集性.例如, 7氯代和8氯代二口恶英和呋喃的营养级放大系数非常低, 原因是这些物质的分子较大, 限制了它们通过细胞膜进入细胞, 并且生物体可以通过排泄作用将它们排出体外[35 ,39 ] .Opperhuizen等[40 ] 也认为疏水性很强的PCDDs和PCDFs在生物体内的生物富集性不高是由于氯代程度高和分子结构较大, 阻止其扩散和吸收, 降低了生物富集性. ...
... 4) POPs 的分子量和亲脂性影响生物富集性.Khairy 等[35 ] 研究了几种有机污染物在河口食物网的分布, 结果显示, 小分子量 PAHs 的脂溶性较低(lgK lip-w <4.1), 生物富集因子(lgBAFs)随着脂溶性(lgK lip-w )的增加而线性增长; 分子量增大, 脂溶性增加(4.1<lgK lip-w <5.4), 生物富集因子(lgBAFs)的增加变缓.原因是分子量较低时, PAHs 在水中的溶解度较大, 且在生物体内的生物转化率较低, 吸收速率远高于清除速率, 因此有明显的生物富集现象.而当分子量增大时, PAHs 的生物转化率提高, 生物富集系数降低. ...
... Physiological features that influence the bioaccumulation
生理特征 对BAFs的影响 主要文献 生长率 生长率加快, 导致“生长稀释”, 降低水柱、浮游生物等生物体内的POPs浓度 [23 ] 性别 雌性生物通过产卵, 降低体内POPs浓度 [23 , 43 -44 ] 脂质 脂质与体内POPs浓度呈正相关 [33 , 45 ] 生物转化 生物转化会降低POPs浓度, 不同生物降解不同种类POPs的能力不同 [35 , 38 -39 ] 体型大小 有研究认为, 体型增大, 由于吸收、储存、转化等因素, 生物体对POPs的生物富集性降低; [46 ,47 ,48 ,49 ,50 ,51 ]
生长率的提高会降低生物体内POPs 浓度.在夏季富营养化的湖泊中, 浮游植物生长最快, 生物量增加, 此时体内 POPs 浓度较低, 尤其是高氯代PCBs 等疏水性较强的化合物.原因在于: 1) 浮游植物大量生长、生物量增大, 形成稀释效应; 2) 浮游植物对 POPs 的大量吸收, 打破了水-气界面的平衡分配, 降低了水柱中溶解态POPs的浓度; 3) 浮游植物对 POPs 的吸收分为两个主要的步骤, 首先是POPs 被吸附在浮游植物的表面(较快), 之后 POPs从细胞的表面转移到细胞内部(较慢), 分子越大, 脂溶性越强, 转移越慢.当浮游植物生长率较快时, POPs 在浮游植物和水柱之间分配失衡.因此浮游植物的生长率越快, 生物量越大, POPs 的脂溶性越强, 生长稀释效应就越大.不仅浮游植物如此, 鱼类等其他生物当生长率大大提高的时候, 其体内的POPs浓度也会下降[23 ] . ...
... POPs 在生物体内的生物转化能够降低其生物富集性.鱼类和很多的水生生物能够降解代谢不同种类的 POPs 物质.例如, 一种吡啶酯的实际生物富集系数是 5, 但根据辛醇-水分配系数(K ow )估算得到的生物富集系数为 678, 原因就是生物转化降低了污染物的浓度[39 ] .另外, 前面也提到, 相同脂溶性(K ow )的 PAHs 的生物富集系数低于 PCBs, 原因就是生物体更容易分解代谢PAHs[35 ,38 ] . ...
Biological pump control of the fate and distribution of hydrophic organic pollutants in water and plankton. Environ-mental Science and
1
2012
... 1) POPs 的亲脂性和卤代程度会影响生物富集作用及变化.Nizzetto 等[36 ] 在意大利南部的马久里湖发现, 春季水华爆发前后, 对于 lgK ow >6.7 (K ow 为辛醇-水分配系数)的 PCBs 来说, 水柱中悬浮颗粒的吸附量占绝大部分, 约为 50%~75%; 对于lgK ow <6.7 的 PCBs 来说, 水柱中溶解态的 PCBs 占优势, 为 5%~35%.在浮游动物体内, 低氯代 PCBs (lgK ow <6.45)的生物富集因子(BAFs)的最大值发生在水华爆发高峰期, 高氯代PCBs (lgK ow >6.45)的生物富集因子(BAFs)最大值发生在水华爆发之前. ...
O’Hara T M, Teixeira C, et al. Spatial trends and bioaccumulation of organochlorine pollu-tants in marine zooplankton from the Alaskan and Canadian Arctic
1
2002
... 2) POPs 的种类、亲脂性和结构会影响其生物集性及其在生物体内的生物转化和代谢分解.Mizukawa 等[3 ] 分析日本东京湾中双壳类、螃蟹、鱼、海水以及沉积物中所含的 20 种 PBDEs 和PCBs 的生物富集性, 发现 PBDEs 和 PCBs 的生物浓缩因子(BCFs)随着 K ow 的增加而增加, 但是当lgK ow >7以后, BCFs 随着 K ow 的增加而下降.PCBs和大多数的 PBDEs 同系物存在生物放大作用, 但另外几种 PBDEs 同系物不存在生物放大作用, 其原因可能是在生物体内被生物转化或代谢分解了.Hoekstra等[37 ] 研究了几类有机污染物在阿拉斯加和加拿大极地地区的桡足类生物体内的生物富集性, 其大小顺序为: 毒杀芬>PCBs>HCHs>DDT> CHLOR >CIBz.毒杀芬和 PCBs 生物富集性之所以较大, 是因为生物体内细胞色素 P-4502B 同工酶对这两类物质的生物转化量较少.另有研究发现, 疏水性相同的PAHs 的生物富集性小于 PCBs, 因为PAHs 在甲壳类和其他无脊椎动物体内较容易被代谢而排出[35 ,38 ] . ...
Biomagnification of organic pollutants in benthic and pelagic marine food chains from the Baltic Sea
3
2008
... 2) POPs 的种类、亲脂性和结构会影响其生物集性及其在生物体内的生物转化和代谢分解.Mizukawa 等[3 ] 分析日本东京湾中双壳类、螃蟹、鱼、海水以及沉积物中所含的 20 种 PBDEs 和PCBs 的生物富集性, 发现 PBDEs 和 PCBs 的生物浓缩因子(BCFs)随着 K ow 的增加而增加, 但是当lgK ow >7以后, BCFs 随着 K ow 的增加而下降.PCBs和大多数的 PBDEs 同系物存在生物放大作用, 但另外几种 PBDEs 同系物不存在生物放大作用, 其原因可能是在生物体内被生物转化或代谢分解了.Hoekstra等[37 ] 研究了几类有机污染物在阿拉斯加和加拿大极地地区的桡足类生物体内的生物富集性, 其大小顺序为: 毒杀芬>PCBs>HCHs>DDT> CHLOR >CIBz.毒杀芬和 PCBs 生物富集性之所以较大, 是因为生物体内细胞色素 P-4502B 同工酶对这两类物质的生物转化量较少.另有研究发现, 疏水性相同的PAHs 的生物富集性小于 PCBs, 因为PAHs 在甲壳类和其他无脊椎动物体内较容易被代谢而排出[35 ,38 ] . ...
... Physiological features that influence the bioaccumulation
生理特征 对BAFs的影响 主要文献 生长率 生长率加快, 导致“生长稀释”, 降低水柱、浮游生物等生物体内的POPs浓度 [23 ] 性别 雌性生物通过产卵, 降低体内POPs浓度 [23 , 43 -44 ] 脂质 脂质与体内POPs浓度呈正相关 [33 , 45 ] 生物转化 生物转化会降低POPs浓度, 不同生物降解不同种类POPs的能力不同 [35 , 38 -39 ] 体型大小 有研究认为, 体型增大, 由于吸收、储存、转化等因素, 生物体对POPs的生物富集性降低; [46 ,47 ,48 ,49 ,50 ,51 ]
生长率的提高会降低生物体内POPs 浓度.在夏季富营养化的湖泊中, 浮游植物生长最快, 生物量增加, 此时体内 POPs 浓度较低, 尤其是高氯代PCBs 等疏水性较强的化合物.原因在于: 1) 浮游植物大量生长、生物量增大, 形成稀释效应; 2) 浮游植物对 POPs 的大量吸收, 打破了水-气界面的平衡分配, 降低了水柱中溶解态POPs的浓度; 3) 浮游植物对 POPs 的吸收分为两个主要的步骤, 首先是POPs 被吸附在浮游植物的表面(较快), 之后 POPs从细胞的表面转移到细胞内部(较慢), 分子越大, 脂溶性越强, 转移越慢.当浮游植物生长率较快时, POPs 在浮游植物和水柱之间分配失衡.因此浮游植物的生长率越快, 生物量越大, POPs 的脂溶性越强, 生长稀释效应就越大.不仅浮游植物如此, 鱼类等其他生物当生长率大大提高的时候, 其体内的POPs浓度也会下降[23 ] . ...
... POPs 在生物体内的生物转化能够降低其生物富集性.鱼类和很多的水生生物能够降解代谢不同种类的 POPs 物质.例如, 一种吡啶酯的实际生物富集系数是 5, 但根据辛醇-水分配系数(K ow )估算得到的生物富集系数为 678, 原因就是生物转化降低了污染物的浓度[39 ] .另外, 前面也提到, 相同脂溶性(K ow )的 PAHs 的生物富集系数低于 PCBs, 原因就是生物体更容易分解代谢PAHs[35 ,38 ] . ...
Bioconcentration. Environmental Scie-nce and
3
1990
... 3) 分子结构和卤代程度会影响 POPs 的生物富集性.例如, 7氯代和8氯代二口恶英和呋喃的营养级放大系数非常低, 原因是这些物质的分子较大, 限制了它们通过细胞膜进入细胞, 并且生物体可以通过排泄作用将它们排出体外[35 ,39 ] .Opperhuizen等[40 ] 也认为疏水性很强的PCDDs和PCDFs在生物体内的生物富集性不高是由于氯代程度高和分子结构较大, 阻止其扩散和吸收, 降低了生物富集性. ...
... Physiological features that influence the bioaccumulation
生理特征 对BAFs的影响 主要文献 生长率 生长率加快, 导致“生长稀释”, 降低水柱、浮游生物等生物体内的POPs浓度 [23 ] 性别 雌性生物通过产卵, 降低体内POPs浓度 [23 , 43 -44 ] 脂质 脂质与体内POPs浓度呈正相关 [33 , 45 ] 生物转化 生物转化会降低POPs浓度, 不同生物降解不同种类POPs的能力不同 [35 , 38 -39 ] 体型大小 有研究认为, 体型增大, 由于吸收、储存、转化等因素, 生物体对POPs的生物富集性降低; [46 ,47 ,48 ,49 ,50 ,51 ]
生长率的提高会降低生物体内POPs 浓度.在夏季富营养化的湖泊中, 浮游植物生长最快, 生物量增加, 此时体内 POPs 浓度较低, 尤其是高氯代PCBs 等疏水性较强的化合物.原因在于: 1) 浮游植物大量生长、生物量增大, 形成稀释效应; 2) 浮游植物对 POPs 的大量吸收, 打破了水-气界面的平衡分配, 降低了水柱中溶解态POPs的浓度; 3) 浮游植物对 POPs 的吸收分为两个主要的步骤, 首先是POPs 被吸附在浮游植物的表面(较快), 之后 POPs从细胞的表面转移到细胞内部(较慢), 分子越大, 脂溶性越强, 转移越慢.当浮游植物生长率较快时, POPs 在浮游植物和水柱之间分配失衡.因此浮游植物的生长率越快, 生物量越大, POPs 的脂溶性越强, 生长稀释效应就越大.不仅浮游植物如此, 鱼类等其他生物当生长率大大提高的时候, 其体内的POPs浓度也会下降[23 ] . ...
... POPs 在生物体内的生物转化能够降低其生物富集性.鱼类和很多的水生生物能够降解代谢不同种类的 POPs 物质.例如, 一种吡啶酯的实际生物富集系数是 5, 但根据辛醇-水分配系数(K ow )估算得到的生物富集系数为 678, 原因就是生物转化降低了污染物的浓度[39 ] .另外, 前面也提到, 相同脂溶性(K ow )的 PAHs 的生物富集系数低于 PCBs, 原因就是生物体更容易分解代谢PAHs[35 ,38 ] . ...
Sijm D T H M. Bioaccumulation and biotransformation of polychlorinated Dibenzo-p-Dioxins and Dibenzofurans in fish
1
1990
... 3) 分子结构和卤代程度会影响 POPs 的生物富集性.例如, 7氯代和8氯代二口恶英和呋喃的营养级放大系数非常低, 原因是这些物质的分子较大, 限制了它们通过细胞膜进入细胞, 并且生物体可以通过排泄作用将它们排出体外[35 ,39 ] .Opperhuizen等[40 ] 也认为疏水性很强的PCDDs和PCDFs在生物体内的生物富集性不高是由于氯代程度高和分子结构较大, 阻止其扩散和吸收, 降低了生物富集性. ...
Bio-accumulation of organochlorines through a remote freshwater food web in the Canadian Arctic
1
1998
... 研究证明, 生物个体的生理特征等因素(如: 生长率、性别、脂含量、生物转化和体型等)会影响POPs的生物富集性[41 ,42 ] (表1 ). ...
Association between perfluorinated compounds and pathological conditions in southern sea otters
1
2006
... 研究证明, 生物个体的生理特征等因素(如: 生长率、性别、脂含量、生物转化和体型等)会影响POPs的生物富集性[41 ,42 ] (表1 ). ...
Factors affecting the elimination of PCBs in the marine copepod Acartiatonsa
2
1983
... Physiological features that influence the bioaccumulation
生理特征 对BAFs的影响 主要文献 生长率 生长率加快, 导致“生长稀释”, 降低水柱、浮游生物等生物体内的POPs浓度 [23 ] 性别 雌性生物通过产卵, 降低体内POPs浓度 [23 , 43 -44 ] 脂质 脂质与体内POPs浓度呈正相关 [33 , 45 ] 生物转化 生物转化会降低POPs浓度, 不同生物降解不同种类POPs的能力不同 [35 , 38 -39 ] 体型大小 有研究认为, 体型增大, 由于吸收、储存、转化等因素, 生物体对POPs的生物富集性降低; [46 ,47 ,48 ,49 ,50 ,51 ]
生长率的提高会降低生物体内POPs 浓度.在夏季富营养化的湖泊中, 浮游植物生长最快, 生物量增加, 此时体内 POPs 浓度较低, 尤其是高氯代PCBs 等疏水性较强的化合物.原因在于: 1) 浮游植物大量生长、生物量增大, 形成稀释效应; 2) 浮游植物对 POPs 的大量吸收, 打破了水-气界面的平衡分配, 降低了水柱中溶解态POPs的浓度; 3) 浮游植物对 POPs 的吸收分为两个主要的步骤, 首先是POPs 被吸附在浮游植物的表面(较快), 之后 POPs从细胞的表面转移到细胞内部(较慢), 分子越大, 脂溶性越强, 转移越慢.当浮游植物生长率较快时, POPs 在浮游植物和水柱之间分配失衡.因此浮游植物的生长率越快, 生物量越大, POPs 的脂溶性越强, 生长稀释效应就越大.不仅浮游植物如此, 鱼类等其他生物当生长率大大提高的时候, 其体内的POPs浓度也会下降[23 ] . ...
... 雄性生物体内的 POPs 浓度高于雌性, 这是因为雌性生物会通过产卵降低体内 POPs 的浓度.研究发现, 雌性梭子鱼体内的 POPs 浓度随着年龄、体重和体型的增大而线性下降, 这是由雌鱼的季节性产卵造成的, 卵中含有的脂质含量比肌肉高10 倍, 其污染物的浓度也相应地高 10 倍左右, 而雄性通过性器官排泄的污染物较少[23 ] .此外, McManus 等[43 ] 发现, 雌性桡足类动物在产卵过程中, 体内的 PCBs 浓度也下降较快, 同样是由于很大一部分 PCBs 转移到富含脂肪的卵上.Lotufo[44 ] 将小型的底栖桡足类暴露在荧蒽中, 发现有 50%的化合物留在卵中, 通过产卵排出体外. ...
Bioaccumulation of sediment-associated fluoranthene in benthic copepods: uptake, elimination and biotransformation
2
1998
... Physiological features that influence the bioaccumulation
生理特征 对BAFs的影响 主要文献 生长率 生长率加快, 导致“生长稀释”, 降低水柱、浮游生物等生物体内的POPs浓度 [23 ] 性别 雌性生物通过产卵, 降低体内POPs浓度 [23 , 43 -44 ] 脂质 脂质与体内POPs浓度呈正相关 [33 , 45 ] 生物转化 生物转化会降低POPs浓度, 不同生物降解不同种类POPs的能力不同 [35 , 38 -39 ] 体型大小 有研究认为, 体型增大, 由于吸收、储存、转化等因素, 生物体对POPs的生物富集性降低; [46 ,47 ,48 ,49 ,50 ,51 ]
生长率的提高会降低生物体内POPs 浓度.在夏季富营养化的湖泊中, 浮游植物生长最快, 生物量增加, 此时体内 POPs 浓度较低, 尤其是高氯代PCBs 等疏水性较强的化合物.原因在于: 1) 浮游植物大量生长、生物量增大, 形成稀释效应; 2) 浮游植物对 POPs 的大量吸收, 打破了水-气界面的平衡分配, 降低了水柱中溶解态POPs的浓度; 3) 浮游植物对 POPs 的吸收分为两个主要的步骤, 首先是POPs 被吸附在浮游植物的表面(较快), 之后 POPs从细胞的表面转移到细胞内部(较慢), 分子越大, 脂溶性越强, 转移越慢.当浮游植物生长率较快时, POPs 在浮游植物和水柱之间分配失衡.因此浮游植物的生长率越快, 生物量越大, POPs 的脂溶性越强, 生长稀释效应就越大.不仅浮游植物如此, 鱼类等其他生物当生长率大大提高的时候, 其体内的POPs浓度也会下降[23 ] . ...
... 雄性生物体内的 POPs 浓度高于雌性, 这是因为雌性生物会通过产卵降低体内 POPs 的浓度.研究发现, 雌性梭子鱼体内的 POPs 浓度随着年龄、体重和体型的增大而线性下降, 这是由雌鱼的季节性产卵造成的, 卵中含有的脂质含量比肌肉高10 倍, 其污染物的浓度也相应地高 10 倍左右, 而雄性通过性器官排泄的污染物较少[23 ] .此外, McManus 等[43 ] 发现, 雌性桡足类动物在产卵过程中, 体内的 PCBs 浓度也下降较快, 同样是由于很大一部分 PCBs 转移到富含脂肪的卵上.Lotufo[44 ] 将小型的底栖桡足类暴露在荧蒽中, 发现有 50%的化合物留在卵中, 通过产卵排出体外. ...
Biomagnification of DDT through the benthic and pelagic food webs of Lake Malawi, East Africa: importance of trophic level and carbon source
2
2001
... Physiological features that influence the bioaccumulation
生理特征 对BAFs的影响 主要文献 生长率 生长率加快, 导致“生长稀释”, 降低水柱、浮游生物等生物体内的POPs浓度 [23 ] 性别 雌性生物通过产卵, 降低体内POPs浓度 [23 , 43 -44 ] 脂质 脂质与体内POPs浓度呈正相关 [33 , 45 ] 生物转化 生物转化会降低POPs浓度, 不同生物降解不同种类POPs的能力不同 [35 , 38 -39 ] 体型大小 有研究认为, 体型增大, 由于吸收、储存、转化等因素, 生物体对POPs的生物富集性降低; [46 ,47 ,48 ,49 ,50 ,51 ]
生长率的提高会降低生物体内POPs 浓度.在夏季富营养化的湖泊中, 浮游植物生长最快, 生物量增加, 此时体内 POPs 浓度较低, 尤其是高氯代PCBs 等疏水性较强的化合物.原因在于: 1) 浮游植物大量生长、生物量增大, 形成稀释效应; 2) 浮游植物对 POPs 的大量吸收, 打破了水-气界面的平衡分配, 降低了水柱中溶解态POPs的浓度; 3) 浮游植物对 POPs 的吸收分为两个主要的步骤, 首先是POPs 被吸附在浮游植物的表面(较快), 之后 POPs从细胞的表面转移到细胞内部(较慢), 分子越大, 脂溶性越强, 转移越慢.当浮游植物生长率较快时, POPs 在浮游植物和水柱之间分配失衡.因此浮游植物的生长率越快, 生物量越大, POPs 的脂溶性越强, 生长稀释效应就越大.不仅浮游植物如此, 鱼类等其他生物当生长率大大提高的时候, 其体内的POPs浓度也会下降[23 ] . ...
... 生物体的脂质含量与其体内的 POPs浓度具有显著的正相关关系.Kidd 等[45 ] 发现 DDT 和 DDE的浓度与生物体脂质含量显著正相关, 且浮游生物要比相同营养级的底栖生物体内的 DDT 浓度高, 推测原因是前者的脂质含量比后者高.另外, 在寡营养湖泊中, 浮游生物、鱼类等生物体内的 POPs浓度比富营养条件下高很多, 除上面提到的生物稀释等因素以外, 寡营养条件下生物体内脂质含量更高也是一个重要原因[33 ] . ...
The prediction of biocon-centration in fish
2
1984
... Physiological features that influence the bioaccumulation
生理特征 对BAFs的影响 主要文献 生长率 生长率加快, 导致“生长稀释”, 降低水柱、浮游生物等生物体内的POPs浓度 [23 ] 性别 雌性生物通过产卵, 降低体内POPs浓度 [23 , 43 -44 ] 脂质 脂质与体内POPs浓度呈正相关 [33 , 45 ] 生物转化 生物转化会降低POPs浓度, 不同生物降解不同种类POPs的能力不同 [35 , 38 -39 ] 体型大小 有研究认为, 体型增大, 由于吸收、储存、转化等因素, 生物体对POPs的生物富集性降低; [46 ,47 ,48 ,49 ,50 ,51 ]
生长率的提高会降低生物体内POPs 浓度.在夏季富营养化的湖泊中, 浮游植物生长最快, 生物量增加, 此时体内 POPs 浓度较低, 尤其是高氯代PCBs 等疏水性较强的化合物.原因在于: 1) 浮游植物大量生长、生物量增大, 形成稀释效应; 2) 浮游植物对 POPs 的大量吸收, 打破了水-气界面的平衡分配, 降低了水柱中溶解态POPs的浓度; 3) 浮游植物对 POPs 的吸收分为两个主要的步骤, 首先是POPs 被吸附在浮游植物的表面(较快), 之后 POPs从细胞的表面转移到细胞内部(较慢), 分子越大, 脂溶性越强, 转移越慢.当浮游植物生长率较快时, POPs 在浮游植物和水柱之间分配失衡.因此浮游植物的生长率越快, 生物量越大, POPs 的脂溶性越强, 生长稀释效应就越大.不仅浮游植物如此, 鱼类等其他生物当生长率大大提高的时候, 其体内的POPs浓度也会下降[23 ] . ...
... POPs 在生物体内的生物富集与体型大小有很大的关系, 但对于不同物种、不同种类的POPs, 其相关关系是不同的.Davies 等[46 ] 认为很多POPs生物富集性的变化和大小主要是由体型造成的.有研究发现, 鱼类体型增大, 脂质含量会增加, 但是其体内邻苯二甲酸酯的生物浓缩因子(BCFs)却下降为原来的 1/25[47 ] .另有研究发现, 鱼类单位生物量含有的DDT含量同样也随着体型的增加而下降[48 ] .这是由生物体的异速增长和生理过程变化造成的, 因为体型较大的鱼体对有机污染物的吸收变慢, 对污染物的储存能力下降[49 ] .Harding 等[50 ] 的研究结果与此相反, 他们发现浮游生物体内单位脂质的PCBs含量不会随着体型大小的变化而变化, 鱼类单位脂质中 PCBs 含量会随着体型的增大而增大.Kucklick 等[51 ] 在苏必利尔湖的研究中发现, 随着体型的增大, 胡瓜鱼单位鲜重的 PCBs 含量从体型最小时的 21 ng/g 增长到体长为 150~199 mm 时的 53 ng/g, 但单位脂质的 PCBs 含量几乎不变, 他们认为污染物浓度随着体型增大而增大是由脂质含量随着体型增大而增加引起的.综上可知, 由于生物体的生物代谢、脂质含量等原因, 使得体型大小在POPs生物富集方面所起的作用可能是不同的. ...
Effect of body size on the uptake and bioconcentration of di-2-ethylhexyl phthalate in rainbow trout
2
1990
... Physiological features that influence the bioaccumulation
生理特征 对BAFs的影响 主要文献 生长率 生长率加快, 导致“生长稀释”, 降低水柱、浮游生物等生物体内的POPs浓度 [23 ] 性别 雌性生物通过产卵, 降低体内POPs浓度 [23 , 43 -44 ] 脂质 脂质与体内POPs浓度呈正相关 [33 , 45 ] 生物转化 生物转化会降低POPs浓度, 不同生物降解不同种类POPs的能力不同 [35 , 38 -39 ] 体型大小 有研究认为, 体型增大, 由于吸收、储存、转化等因素, 生物体对POPs的生物富集性降低; [46 ,47 ,48 ,49 ,50 ,51 ]
生长率的提高会降低生物体内POPs 浓度.在夏季富营养化的湖泊中, 浮游植物生长最快, 生物量增加, 此时体内 POPs 浓度较低, 尤其是高氯代PCBs 等疏水性较强的化合物.原因在于: 1) 浮游植物大量生长、生物量增大, 形成稀释效应; 2) 浮游植物对 POPs 的大量吸收, 打破了水-气界面的平衡分配, 降低了水柱中溶解态POPs的浓度; 3) 浮游植物对 POPs 的吸收分为两个主要的步骤, 首先是POPs 被吸附在浮游植物的表面(较快), 之后 POPs从细胞的表面转移到细胞内部(较慢), 分子越大, 脂溶性越强, 转移越慢.当浮游植物生长率较快时, POPs 在浮游植物和水柱之间分配失衡.因此浮游植物的生长率越快, 生物量越大, POPs 的脂溶性越强, 生长稀释效应就越大.不仅浮游植物如此, 鱼类等其他生物当生长率大大提高的时候, 其体内的POPs浓度也会下降[23 ] . ...
... POPs 在生物体内的生物富集与体型大小有很大的关系, 但对于不同物种、不同种类的POPs, 其相关关系是不同的.Davies 等[46 ] 认为很多POPs生物富集性的变化和大小主要是由体型造成的.有研究发现, 鱼类体型增大, 脂质含量会增加, 但是其体内邻苯二甲酸酯的生物浓缩因子(BCFs)却下降为原来的 1/25[47 ] .另有研究发现, 鱼类单位生物量含有的DDT含量同样也随着体型的增加而下降[48 ] .这是由生物体的异速增长和生理过程变化造成的, 因为体型较大的鱼体对有机污染物的吸收变慢, 对污染物的储存能力下降[49 ] .Harding 等[50 ] 的研究结果与此相反, 他们发现浮游生物体内单位脂质的PCBs含量不会随着体型大小的变化而变化, 鱼类单位脂质中 PCBs 含量会随着体型的增大而增大.Kucklick 等[51 ] 在苏必利尔湖的研究中发现, 随着体型的增大, 胡瓜鱼单位鲜重的 PCBs 含量从体型最小时的 21 ng/g 增长到体长为 150~199 mm 时的 53 ng/g, 但单位脂质的 PCBs 含量几乎不变, 他们认为污染物浓度随着体型增大而增大是由脂质含量随着体型增大而增加引起的.综上可知, 由于生物体的生物代谢、脂质含量等原因, 使得体型大小在POPs生物富集方面所起的作用可能是不同的. ...
Correlations between respiration and direct uptake of DDT in the mosquito fish Gambusiaaffinis. Bulletin of Environmental Con-tamination and
2
1971
... Physiological features that influence the bioaccumulation
生理特征 对BAFs的影响 主要文献 生长率 生长率加快, 导致“生长稀释”, 降低水柱、浮游生物等生物体内的POPs浓度 [23 ] 性别 雌性生物通过产卵, 降低体内POPs浓度 [23 , 43 -44 ] 脂质 脂质与体内POPs浓度呈正相关 [33 , 45 ] 生物转化 生物转化会降低POPs浓度, 不同生物降解不同种类POPs的能力不同 [35 , 38 -39 ] 体型大小 有研究认为, 体型增大, 由于吸收、储存、转化等因素, 生物体对POPs的生物富集性降低; [46 ,47 ,48 ,49 ,50 ,51 ]
生长率的提高会降低生物体内POPs 浓度.在夏季富营养化的湖泊中, 浮游植物生长最快, 生物量增加, 此时体内 POPs 浓度较低, 尤其是高氯代PCBs 等疏水性较强的化合物.原因在于: 1) 浮游植物大量生长、生物量增大, 形成稀释效应; 2) 浮游植物对 POPs 的大量吸收, 打破了水-气界面的平衡分配, 降低了水柱中溶解态POPs的浓度; 3) 浮游植物对 POPs 的吸收分为两个主要的步骤, 首先是POPs 被吸附在浮游植物的表面(较快), 之后 POPs从细胞的表面转移到细胞内部(较慢), 分子越大, 脂溶性越强, 转移越慢.当浮游植物生长率较快时, POPs 在浮游植物和水柱之间分配失衡.因此浮游植物的生长率越快, 生物量越大, POPs 的脂溶性越强, 生长稀释效应就越大.不仅浮游植物如此, 鱼类等其他生物当生长率大大提高的时候, 其体内的POPs浓度也会下降[23 ] . ...
... POPs 在生物体内的生物富集与体型大小有很大的关系, 但对于不同物种、不同种类的POPs, 其相关关系是不同的.Davies 等[46 ] 认为很多POPs生物富集性的变化和大小主要是由体型造成的.有研究发现, 鱼类体型增大, 脂质含量会增加, 但是其体内邻苯二甲酸酯的生物浓缩因子(BCFs)却下降为原来的 1/25[47 ] .另有研究发现, 鱼类单位生物量含有的DDT含量同样也随着体型的增加而下降[48 ] .这是由生物体的异速增长和生理过程变化造成的, 因为体型较大的鱼体对有机污染物的吸收变慢, 对污染物的储存能力下降[49 ] .Harding 等[50 ] 的研究结果与此相反, 他们发现浮游生物体内单位脂质的PCBs含量不会随着体型大小的变化而变化, 鱼类单位脂质中 PCBs 含量会随着体型的增大而增大.Kucklick 等[51 ] 在苏必利尔湖的研究中发现, 随着体型的增大, 胡瓜鱼单位鲜重的 PCBs 含量从体型最小时的 21 ng/g 增长到体长为 150~199 mm 时的 53 ng/g, 但单位脂质的 PCBs 含量几乎不变, 他们认为污染物浓度随着体型增大而增大是由脂质含量随着体型增大而增加引起的.综上可知, 由于生物体的生物代谢、脂质含量等原因, 使得体型大小在POPs生物富集方面所起的作用可能是不同的. ...
Quantitative relations in the physiologi-cal constitutions of mammals
2
1949
... Physiological features that influence the bioaccumulation
生理特征 对BAFs的影响 主要文献 生长率 生长率加快, 导致“生长稀释”, 降低水柱、浮游生物等生物体内的POPs浓度 [23 ] 性别 雌性生物通过产卵, 降低体内POPs浓度 [23 , 43 -44 ] 脂质 脂质与体内POPs浓度呈正相关 [33 , 45 ] 生物转化 生物转化会降低POPs浓度, 不同生物降解不同种类POPs的能力不同 [35 , 38 -39 ] 体型大小 有研究认为, 体型增大, 由于吸收、储存、转化等因素, 生物体对POPs的生物富集性降低; [46 ,47 ,48 ,49 ,50 ,51 ]
生长率的提高会降低生物体内POPs 浓度.在夏季富营养化的湖泊中, 浮游植物生长最快, 生物量增加, 此时体内 POPs 浓度较低, 尤其是高氯代PCBs 等疏水性较强的化合物.原因在于: 1) 浮游植物大量生长、生物量增大, 形成稀释效应; 2) 浮游植物对 POPs 的大量吸收, 打破了水-气界面的平衡分配, 降低了水柱中溶解态POPs的浓度; 3) 浮游植物对 POPs 的吸收分为两个主要的步骤, 首先是POPs 被吸附在浮游植物的表面(较快), 之后 POPs从细胞的表面转移到细胞内部(较慢), 分子越大, 脂溶性越强, 转移越慢.当浮游植物生长率较快时, POPs 在浮游植物和水柱之间分配失衡.因此浮游植物的生长率越快, 生物量越大, POPs 的脂溶性越强, 生长稀释效应就越大.不仅浮游植物如此, 鱼类等其他生物当生长率大大提高的时候, 其体内的POPs浓度也会下降[23 ] . ...
... POPs 在生物体内的生物富集与体型大小有很大的关系, 但对于不同物种、不同种类的POPs, 其相关关系是不同的.Davies 等[46 ] 认为很多POPs生物富集性的变化和大小主要是由体型造成的.有研究发现, 鱼类体型增大, 脂质含量会增加, 但是其体内邻苯二甲酸酯的生物浓缩因子(BCFs)却下降为原来的 1/25[47 ] .另有研究发现, 鱼类单位生物量含有的DDT含量同样也随着体型的增加而下降[48 ] .这是由生物体的异速增长和生理过程变化造成的, 因为体型较大的鱼体对有机污染物的吸收变慢, 对污染物的储存能力下降[49 ] .Harding 等[50 ] 的研究结果与此相反, 他们发现浮游生物体内单位脂质的PCBs含量不会随着体型大小的变化而变化, 鱼类单位脂质中 PCBs 含量会随着体型的增大而增大.Kucklick 等[51 ] 在苏必利尔湖的研究中发现, 随着体型的增大, 胡瓜鱼单位鲜重的 PCBs 含量从体型最小时的 21 ng/g 增长到体长为 150~199 mm 时的 53 ng/g, 但单位脂质的 PCBs 含量几乎不变, 他们认为污染物浓度随着体型增大而增大是由脂质含量随着体型增大而增加引起的.综上可知, 由于生物体的生物代谢、脂质含量等原因, 使得体型大小在POPs生物富集方面所起的作用可能是不同的. ...
LeBlanc R J, Wass W P, et al. Bioaccumulation of polychlorinated biphenyls (PCBs) in the marine pelagic food web, based on a seasonal study in the southern Gulf of St. Lawrence, 1976-1977
2
1997
... Physiological features that influence the bioaccumulation
生理特征 对BAFs的影响 主要文献 生长率 生长率加快, 导致“生长稀释”, 降低水柱、浮游生物等生物体内的POPs浓度 [23 ] 性别 雌性生物通过产卵, 降低体内POPs浓度 [23 , 43 -44 ] 脂质 脂质与体内POPs浓度呈正相关 [33 , 45 ] 生物转化 生物转化会降低POPs浓度, 不同生物降解不同种类POPs的能力不同 [35 , 38 -39 ] 体型大小 有研究认为, 体型增大, 由于吸收、储存、转化等因素, 生物体对POPs的生物富集性降低; [46 ,47 ,48 ,49 ,50 ,51 ]
生长率的提高会降低生物体内POPs 浓度.在夏季富营养化的湖泊中, 浮游植物生长最快, 生物量增加, 此时体内 POPs 浓度较低, 尤其是高氯代PCBs 等疏水性较强的化合物.原因在于: 1) 浮游植物大量生长、生物量增大, 形成稀释效应; 2) 浮游植物对 POPs 的大量吸收, 打破了水-气界面的平衡分配, 降低了水柱中溶解态POPs的浓度; 3) 浮游植物对 POPs 的吸收分为两个主要的步骤, 首先是POPs 被吸附在浮游植物的表面(较快), 之后 POPs从细胞的表面转移到细胞内部(较慢), 分子越大, 脂溶性越强, 转移越慢.当浮游植物生长率较快时, POPs 在浮游植物和水柱之间分配失衡.因此浮游植物的生长率越快, 生物量越大, POPs 的脂溶性越强, 生长稀释效应就越大.不仅浮游植物如此, 鱼类等其他生物当生长率大大提高的时候, 其体内的POPs浓度也会下降[23 ] . ...
... POPs 在生物体内的生物富集与体型大小有很大的关系, 但对于不同物种、不同种类的POPs, 其相关关系是不同的.Davies 等[46 ] 认为很多POPs生物富集性的变化和大小主要是由体型造成的.有研究发现, 鱼类体型增大, 脂质含量会增加, 但是其体内邻苯二甲酸酯的生物浓缩因子(BCFs)却下降为原来的 1/25[47 ] .另有研究发现, 鱼类单位生物量含有的DDT含量同样也随着体型的增加而下降[48 ] .这是由生物体的异速增长和生理过程变化造成的, 因为体型较大的鱼体对有机污染物的吸收变慢, 对污染物的储存能力下降[49 ] .Harding 等[50 ] 的研究结果与此相反, 他们发现浮游生物体内单位脂质的PCBs含量不会随着体型大小的变化而变化, 鱼类单位脂质中 PCBs 含量会随着体型的增大而增大.Kucklick 等[51 ] 在苏必利尔湖的研究中发现, 随着体型的增大, 胡瓜鱼单位鲜重的 PCBs 含量从体型最小时的 21 ng/g 增长到体长为 150~199 mm 时的 53 ng/g, 但单位脂质的 PCBs 含量几乎不变, 他们认为污染物浓度随着体型增大而增大是由脂质含量随着体型增大而增加引起的.综上可知, 由于生物体的生物代谢、脂质含量等原因, 使得体型大小在POPs生物富集方面所起的作用可能是不同的. ...
Organochlorines in Lake Superior’s food web
2
1998
... Physiological features that influence the bioaccumulation
生理特征 对BAFs的影响 主要文献 生长率 生长率加快, 导致“生长稀释”, 降低水柱、浮游生物等生物体内的POPs浓度 [23 ] 性别 雌性生物通过产卵, 降低体内POPs浓度 [23 , 43 -44 ] 脂质 脂质与体内POPs浓度呈正相关 [33 , 45 ] 生物转化 生物转化会降低POPs浓度, 不同生物降解不同种类POPs的能力不同 [35 , 38 -39 ] 体型大小 有研究认为, 体型增大, 由于吸收、储存、转化等因素, 生物体对POPs的生物富集性降低; [46 ,47 ,48 ,49 ,50 ,51 ]
生长率的提高会降低生物体内POPs 浓度.在夏季富营养化的湖泊中, 浮游植物生长最快, 生物量增加, 此时体内 POPs 浓度较低, 尤其是高氯代PCBs 等疏水性较强的化合物.原因在于: 1) 浮游植物大量生长、生物量增大, 形成稀释效应; 2) 浮游植物对 POPs 的大量吸收, 打破了水-气界面的平衡分配, 降低了水柱中溶解态POPs的浓度; 3) 浮游植物对 POPs 的吸收分为两个主要的步骤, 首先是POPs 被吸附在浮游植物的表面(较快), 之后 POPs从细胞的表面转移到细胞内部(较慢), 分子越大, 脂溶性越强, 转移越慢.当浮游植物生长率较快时, POPs 在浮游植物和水柱之间分配失衡.因此浮游植物的生长率越快, 生物量越大, POPs 的脂溶性越强, 生长稀释效应就越大.不仅浮游植物如此, 鱼类等其他生物当生长率大大提高的时候, 其体内的POPs浓度也会下降[23 ] . ...
... POPs 在生物体内的生物富集与体型大小有很大的关系, 但对于不同物种、不同种类的POPs, 其相关关系是不同的.Davies 等[46 ] 认为很多POPs生物富集性的变化和大小主要是由体型造成的.有研究发现, 鱼类体型增大, 脂质含量会增加, 但是其体内邻苯二甲酸酯的生物浓缩因子(BCFs)却下降为原来的 1/25[47 ] .另有研究发现, 鱼类单位生物量含有的DDT含量同样也随着体型的增加而下降[48 ] .这是由生物体的异速增长和生理过程变化造成的, 因为体型较大的鱼体对有机污染物的吸收变慢, 对污染物的储存能力下降[49 ] .Harding 等[50 ] 的研究结果与此相反, 他们发现浮游生物体内单位脂质的PCBs含量不会随着体型大小的变化而变化, 鱼类单位脂质中 PCBs 含量会随着体型的增大而增大.Kucklick 等[51 ] 在苏必利尔湖的研究中发现, 随着体型的增大, 胡瓜鱼单位鲜重的 PCBs 含量从体型最小时的 21 ng/g 增长到体长为 150~199 mm 时的 53 ng/g, 但单位脂质的 PCBs 含量几乎不变, 他们认为污染物浓度随着体型增大而增大是由脂质含量随着体型增大而增加引起的.综上可知, 由于生物体的生物代谢、脂质含量等原因, 使得体型大小在POPs生物富集方面所起的作用可能是不同的. ...
〈
〉



{kind=link}
{kind=link}
{kind=link}
{kind=link}