
王秋懿, 郭伟超, 王韬, 郝倩, 许重阳, 柳絮, 韩玥, 邱爽, 刘鸿雁
北京大学城市与环境学院, 北京 100871
Qiuyi WANG, Weichao GUO, Tao WANG, Qian HAO, Chongyang XU, Xu LIU, Yue HAN, Shuang QIU, Hongyan LIU
通讯作者:
收稿日期:
2016-02-29
修回日期:
2016-04-25
网络出版日期:
2017-05-11
版权声明:
2017 《北京大学学报(自然科学版)》编辑部 《北京大学学报(自然科学版)》编辑部 所有
基金资助:
展开
摘要 根据京津冀山地460个样地调查的结果, 对该地区有刺灌木的分布格局及影响因子进行分析。结果表明, 有刺灌木在约60%的样地出现, 主要为荆条灌丛样地, 在少数样地甚至成为灌木层优势种。有刺灌木共24种, 主要来自鼠李科、蔷薇科和豆科。恒有度最高的3种有刺灌木(酸枣、小叶鼠李和红花锦鸡儿)主要分布于海拔600 m以下的低山丘陵区较缓的阳坡和半阳坡。定量分析结果表明, 光照和干旱是限制常见有刺灌木(尤其是酸枣)生长的重要因子。受气候、地形和群落因素的共同影响, 有刺灌木在京津冀山地不同生境条件下均有分布。
关键词:
Abstract To analyze the distribution pattern of thorny shrubs and its impact factors, 460 sample sites were collected in mountains of the Beijing-Tianjin-Hebei region. Results showed that about 60% of sample sites had thorny shrubs, which were mainly the shrub community dominated by Vitex negundo var. heterophylla. In a small number of sample sites, thorny shrubs even became the dominant species. Twenty-four thorny shrub species were mainly from the families Rhamnaceae, Rosaceae and Leguminosae. The most common three species (Ziziphus jujuba, Rhamnus parvifolia and Caragana rosea) are mainly distributed on the gentle sunny and half-sunny slopes in the hilly area with elevation lower than 600 m a.s.l.. Quantitative analysis also showed that light condition and drought were limiting factors for growth of thorny shrubs (especially Ziziphus jujuba). Affected by climate, topography, and community factors together, thorny shrubs can be distributed under different habitats in mountains of the Beijing-Tianjin-Hebei region.
Keywords:
灌木在世界上分布广泛, 具有很强的环境适应性, 能够通过多样化的形态结构、生理过程和功能属性, 耐受或规避干旱胁迫, 在干旱、半干旱生态系统中十分常见[1,2,3,4]。植株多刺通常是灌木适应干旱的特征之一。刺状的叶片、枝条有利于减少太阳辐射面积和保存植物体内水分。研究表明, 在降水稀少、岩石裸露、土壤贫瘠的地区容易出现矮化、多刺的旱生灌木类型, 其种类丰富度和多样性也与干旱程度密切相关[1,5-7]。此外, 植株多刺能够防止食草动物大量啃食, 对自身和周边物种均可起到一定的保护作用, 因而在一些地区, 多刺灌丛表现为荒地、草原向郁闭森林过渡的中间形态[1,2,3,4]。在我国干旱半干旱地区, 土地荒漠化的加剧使灌木的分布范围不断扩大, 遭到破坏的森林植被被次生灌木取代, 部分有刺灌木则作为绿化荒山的先锋树种引种栽培, 成为我国生态恢复建设的重要资源和理想材料[4,11-12]。因此, 探究有刺灌木的分布格局有助于深入理解其与环境条件的作用关系, 对分析未来气候变化和土地利用变化中灌丛群落的响应方式有重要意义。
京津冀山地分布着大量灌丛群落, 森林中也常见茂密灌木层。该地区复杂的环境条件和长期的人为活动历史使得灌木种类逐渐在中低山自然植被中占据优势地位, 土壤环境因素和地形因素对灌木分布格局影响较大[13,14]。然而, 以往的研究对当地广泛存在的有刺灌木类型关注较少, 对环境因子与有刺灌木分布格局形成的关系缺乏认识。本文采用野外植被调查手段, 选取研究区内 460 个样地数据, 对该地区有刺灌木的分布格局进行考察, 并结合各样方的地形、气候等环境指标, 分析形成该种格局的影响因子。
京津冀山地主要由太行山脉和燕山山脉构成, 太行山脉位于河北、山西、河南三省之间, 呈东北-西南走向, 地势北高南低, 海拔总体上在 2000 m以下。燕山山脉处于北京与河北的北部, 东西方向较长。两山脉在北京西部相接, 呈弧状包围河北平 原[15,16]。
该地区属于温带半湿润大陆性季风气候, 向西北部半干旱区过渡, 年均温在-1~15 °C 之间, 年降水在 400~700 mm 之间, 季节性明显, 雨热同期。年均温南北差异大, 山区的复杂地形对局地气温有明显影响。降水量空间分布不均匀,受地形影响, 山地迎风坡为多雨地带, 年降水可达 800 mm, 山间盆地则降水较少[15,16]。
该地区植物种类丰富, 以北温带区系成分占绝对优势, 起源于北极第三纪植物区系,同时受周边欧洲西伯利亚成分、东亚成分、热带成分等影响, 具有较大的过渡性特点, 大量具有热带、亚热带亲缘的科属向北分布, 在该地区普遍存在, 由历史原因和地理气候条件共同形成当前的分布格局[17,18]。当地自然植被类型一般应为落叶阔叶林, 但由于长期人为活动的干扰, 原始植被已经基本上被破坏, 现存的仅为次生植被[19]。太行山东麓、燕山南麓海拔 1000 m 以下的低山丘陵区主要为荆条(Vitex negundo var. heterophylla)、酸枣(Ziziphus jujuba)和黄栌(Cotinus coggygria)组成的灌丛和白羊草(Bothriochloa ischaemum)-黄背草(Themeda japonica)草坡, 海拔 800~1300 m 的广大山地分布有虎榛子(Ostryopsis davidiana)-绣线菊(Spiraea salicifolia)灌丛, 均属于森林破坏后的次生植被类型。中北部中山区有次生的落叶阔叶林, 海拔 1200 m 以下多为辽东栎林(Quercus wutaishanica), 混生有油松林(Pinus tabuliformis); 海拔1200~1800 m主要为白桦(Betula platyphylla)、山杨(Populus davidiana)等落叶阔叶林; 中山顶部 1800 m 以上有青扦(Picea wilsonii)、白扦(Picea meyeri)、华北落叶松(Larix principis-rupprechtii)组成的针叶林; 海拔 2500 m以上的山地顶部则为亚高山草甸[19,20]。
研究区土壤以褐土分布最广, 其范围与山地丘陵带灌丛、灌草丛植被分布基本上一致; 中山上部有小面积棕壤和山地草甸土, 与森林植被和亚高山草甸分布基本上一致; 山地北缘栗钙土和暗栗钙土地区主要为羊草草原和杂类草草原[19,20]。
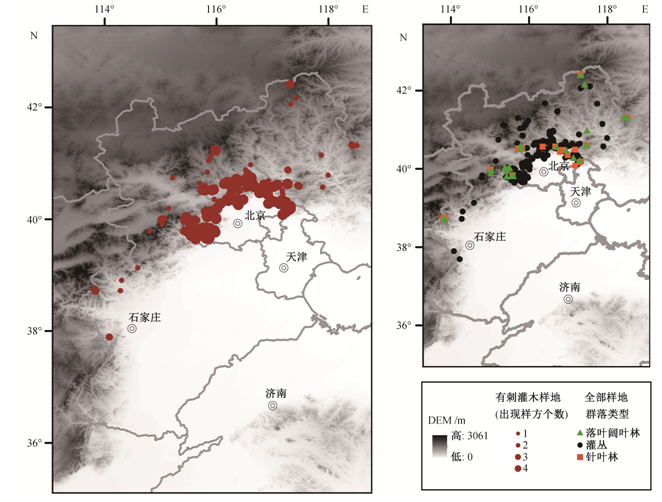
本研究组于 2011—2014 年在研究区选取 460个具有代表性的样地进行植被调查, 其中包括 79个落叶阔叶林样地、47 个针叶林样地和 334 个灌丛样地(图 1), 记录每个样地的经纬度、海拔、坡度和坡向信息。森林样地的面积为 20 m×50 m 或20 m×30 m, 在每个样地内调查 6 个 5 m×5 m 的灌木样方; 灌丛样地的面积为 10 m×10 m, 在每个样地内调查4个5 m×5 m的灌木样方, 记录样方内每种灌木的多度(或株数)、高度和盖度, 按多度分级记录的样方数据在后续分析中统一量化为株数结果(cop3≈100 株, cop2≈75 株, cop1≈50 株, sp≈25 株, sol≈10株, un≈1株)。
选取已经得到广泛应用的WorldClim气候数据集[21]中多年平均气温和多年平均降水量两项气候指标。该数据集的空间分辨率为 1 km, 时间范围为1950—2000年。依据每个样地的地理位置, 从中提取样地的气候信息进行后续分析。
应用 R 软件的 stats 软件包, 对各有刺灌木种类的属性特征和环境因子信息进行方差分析, 比较各种类间的差别并进行显著性检验。应用 corpcor 和 ggm 软件包对环境因子和有刺灌木属性的关系进行偏相关分析, 给出显著性检验结果。仅在极少数样地出现的有刺灌木种类在统计分析过程中没有计入(表1)。
表1 有刺灌木科属信息统计
Table 1 List of all thorny shrub species in the study area
在全部 460 个样地中, 记录了 1462 个有灌木存在的样方, 包含 103 种灌木。其中, 有刺灌木共24 种, 出现在 274 个样地 640 个样方内, 但仅在其中 32 个样地作为灌木层优势种出现(表 1, 图 2)。有刺灌木种类较多的主要为鼠李科(8 种)、蔷薇科(6 种)和豆科(5 种)。在样方中出现次数最多的有刺灌木依次为酸枣(362 个)、小叶鼠李(183 个)和红花锦鸡儿(85 个), 其中酸枣出现的样方占全部有刺灌木样方的50%以上(表1)。
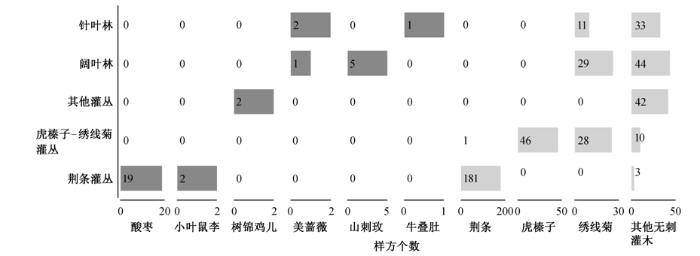
有刺灌木最常见的群落类型为荆条灌丛, 共160个样地, 其中19个样地为酸枣单独或与荆条共同作为优势种, 另有 138 个样地有酸枣伴生, 小叶鼠李则出现在 75 个荆条灌丛样地内, 在其中 2 个样地与荆条共同作为优势种, 在以山杏、绣线菊为优势种的灌丛内也常出现小叶鼠李。共 47 个落叶阔叶林样地和 24 个针叶林样地出现有刺灌木, 以美蔷薇、山刺玫为主, 但仅极个别样地以它们作为灌木层优势种(图2)。
图2 不同群落类型中灌木层优势种信息统计(深灰色表示有刺灌木, 浅灰色表示无刺灌木, 每个数字表示纵轴某类群落中出现横轴上灌木的样方个数) Fig. 2 Summary of dominant shrub species in plant communities in the study area
研究区内有刺灌木总体平均株高为1.12 m, 最高的单个样方株高记录为酸枣, 可达 5.8 m。方差分析结果表明, 不同有刺灌木类型之间平均株高存在显著差异(p<0.001)。平均株高最高的种类是东北鼠李, 为 1.53 m。除小叶鼠李外, 鼠李属各种株高均较高, 平均为 1.03 m, 枣属酸枣的平均株高也较高, 为 1.24 m。株高最低的是小檗属(掌刺小檗), 仅0.50 m (表2)。
全部计入统计的样方内共 11330 株有刺灌木, 平均每个样方约 18 株。不同灌木类型的样方内总株数同样存在显著差别(p<0.001), 总株数最多的有刺灌木是酸枣, 共5493株, 平均每个样方约15株, 鼠李属、锦鸡儿属和蔷薇属总株数均较多, 分别为2715, 1498 和 1085 株, 总株数最少种类的是锦鸡儿, 仅13株, 平均每个样方约3株(表2)。
有刺灌木在各样方的总体平均盖度为 5.16%, 不同物种间存在显著差别(p<0.001)。样方内平均盖度最大的有刺灌木是树锦鸡儿, 达 24.73%, 其单个样方盖度最高为 65%, 是全部记录中有刺灌木盖度最大的。沙棘属和悬钩子属的平均盖度较大, 分别为 8.63%和 6.71%。样方内平均盖度最小的是小檗属(掌刺小檗), 仅0.71% (表2)。
表2 有刺灌木在群落中的重要性指标统计
Table 2 Statistics of community variables of thorny shrub species
从地理分布来看, 研究区内有刺灌木出现在绝大部分调查地区内, 仅燕山北缘小部分样地未见有刺灌木, 在北京周边山区中出现较多, 样地内大部分样方均有出现, 而在河北样地中, 多为 1 个样地内仅1~2个样方可见有刺灌木(图1)。
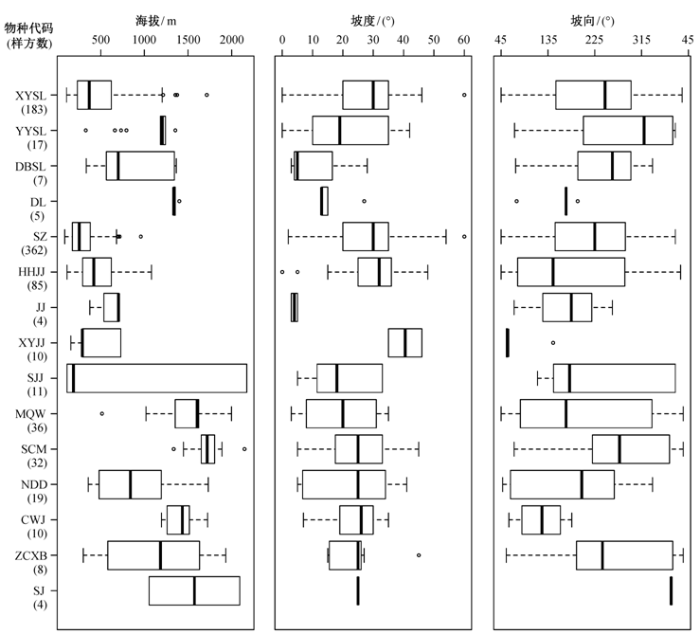
有刺灌木总体分布在海拔 87~2172 m 的山地, 平均海拔高度为 557.3 m, 比其他灌木的分布海拔低(表 3)。各物种间海拔分布区别较大, 存在显著差异(p<0.001)。平均分布海拔最高的是山刺玫, 为1707.5 m, 最低的是酸枣, 为307.6 m。分布海拔范围最广的是树锦鸡儿, 其样方的海拔高差可达 2057 m, 而分布范围最窄的是冻绿, 海拔高差仅66 m。主要分布在1000 m以上中山区的种类有圆叶鼠李、冻绿、美蔷薇、山刺玫、刺五加和沙棘, 主要分布在600 m 以下低山丘陵地带的种类为小叶鼠李、酸枣、红花锦鸡儿和小叶锦鸡儿(图3)。
图3 有刺灌木的海拔、坡度、坡向分布(各物种代码对应名称见 Fig. 3 Altitude, slope and aspect of thorny shrub species
存在有刺灌木的样方总体上位于30°以内的山坡上, 平均坡度为 27.0°, 而全部灌木样方的平均坡度为 39.6°, 说明有刺灌木相对其他灌木所处坡度较缓(表 3)。不同物种间总体差异显著(p<0.001)。主要分布在较陡坡上的有刺灌木只有小叶锦鸡儿, 其平均坡度为 40.5°, 而东北鼠李、冻绿和锦鸡儿则主要分布在 15°以下的缓坡上, 其他种类分布在15~30°范围之间(图3)。
有刺灌木在各个坡向的样方中均出现, 不同种类的坡向分布存在显著差别(p<0.001)。主要分布在阳坡、半阳坡的有刺灌木为冻绿、红花锦鸡儿、锦鸡儿、小叶锦鸡儿、牛叠肚和刺五加, 圆叶鼠李、山刺玫、掌刺小檗和沙棘主要出现在阴坡及半阴坡, 其他种类主要分布在阳坡和半阴坡(图3)。
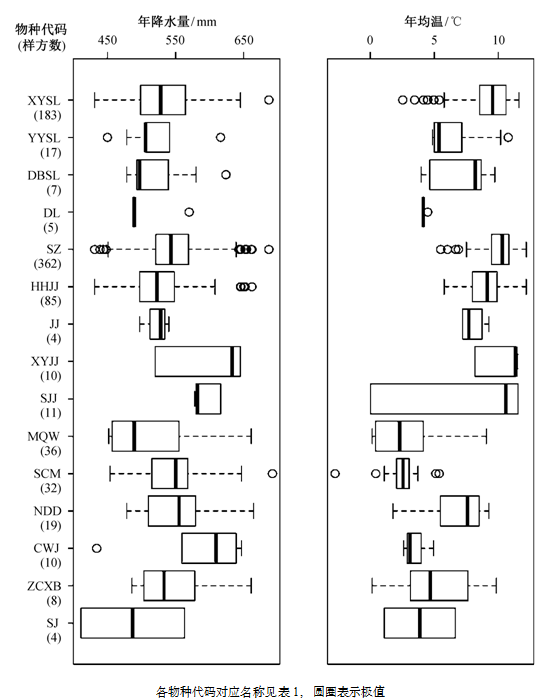
从气候条件来看, 有刺灌木分布在年降水量411~692 mm 的范围内, 与全部灌木样方的年降水量范围基本上一致, 但从均值来看, 有刺灌木样方的平均年降水量为 542.36 mm, 略高于全部灌木样方的结果(表 3)。不同有刺灌木种类之间年降水量存在显著差异(p<0.001)。年降水量最高的为树锦鸡儿, 其均值达593 mm, 各样方降水量均在570 mm以上, 最低的为沙棘, 均值仅 487 mm。主要分布在年降水量大于 550 mm 地区的种类有小叶锦鸡儿、树锦鸡儿和刺五加, 其他种类多分布在 450~ 550 mm范围之间(图4)。
有刺灌木分布的年均温范围为-2.7~12.2°C, 与全部灌木样方的年均温范围相近, 有刺灌木样方的总体均值为 8.7°C, 高于全部灌木样方的结果(表3)。不同种类的有刺灌木年均温存在显著差异(p< 0.001)。年均温最高的是小叶锦鸡儿和酸枣, 均值达 10.1°C; 最低的是美蔷薇, 仅 2.5°C。小叶鼠李、酸枣、红花锦鸡儿和小叶锦鸡儿主要分布在年均温超过 8°C 的地区, 气候相对温暖; 而美蔷薇、山刺玫和刺五加则主要分布在年均温低于5°C的地区(图4)。
图4 有刺灌木的气候因子分布 Fig. 4 Mean annual precipitation and mean annual temperature of thorny shrub species
表3 灌木样方地理环境分布信息统计
Table 3 Environmental factors of shrub sample sites in the study area
偏相关分析结果表明, 有刺灌木整体的株高和盖度都与坡向、年降水和年均温显著正相关, 与坡度关系不显著; 株数仅与年降水显著正相关, 与坡度、坡向和年均温没有显著关系(表4)。
表4 有刺灌木在群落中的重要性指标与环境因子关系的偏相关分析结果
Table 4 Partial correlation of environmental factors and community variables of thorny shrub species
在属的水平上, 恒有度最高的 3 个属株高、株数和盖度变化与环境因子的关系表现不尽相同。枣属(酸枣)表现出株高与年降水显著正相关, 株数与坡向显著负相关, 年均温与株高显著正相关, 与株数显著负相关, 盖度指标与4个环境因子均没有显著相关关系。鼠李属的株高与年均温表现出显著负相关关系, 其他指标与环境因子关系不显著。锦鸡儿属的株高与环境因子均没有显著的相关关系, 其株数和盖度都与坡向、年降水显著正相关, 其中盖度与年降水的相关性较强, 盖度与年均温则存在显著负相关关系(表4)。
植被调查结果表明, 有刺灌木在京津冀山地植被中较为常见, 60%的样地内均存在有刺灌木, 仅在燕山北缘小部分样地未见(图 1)。在全部 105 种灌木中, 有刺灌木共 24 种, 分别来自 6 科 10 属, 鼠李科、蔷薇科和豆科种类较多。有刺灌木在灌丛群落(尤其是荆条灌丛)中大量出现, 占其样地比例的 78%, 并在 10%的样地中(21 个)中作为灌木层优势种; 在 59%的落叶阔叶林和 51%的针叶林样地中也出现有刺灌木(图2)。
从植被区系分析, 研究区有刺灌木多属于温带成分, 其中蔷薇属、小檗属为北温带分布属, 锦鸡儿属为温带亚洲分布属, 沙棘属为旧大陆温带分布属; 鼠李属为世界广布成分; 而枣属为泛热带成分, 五加属为东亚成分, 具有热带亲缘[22]。
从地理分布格局来看, 有刺灌木与其他灌木总体分布的经纬度、海拔范围基本上重合(图 1、表 3), 但有刺灌木平均分布海拔相对较低, 尤其是最常见的 3 个种类(酸枣、小叶鼠李和红花锦鸡儿)均主要分布在 600 m 以下的低山丘陵区(图 3), 这与华北地区荆条优势灌丛多分布于低山地带有关[20], 分布在中山区的虎榛子-绣线菊灌丛中较少出现有刺灌木, 并且没有以有刺灌木为优势种的样方(图2)。
从地形条件来看,相对于其他灌木, 有刺灌木主要分布在坡度较缓的阳坡、半阳坡和半阴坡, 仅有沙棘一种只在阴坡出现(图 3)。以往的研究也表明多数有刺灌木具有喜光的习性, 生长在向阳、干燥的地区[11,23-24]。
从气候条件来看, 有刺灌木分布的降水和温度范围与其他灌木基本上一致, 但年降水量均值和年均温都相对较高。其中小叶锦鸡儿、树锦鸡儿和刺五加多分布在降水条件较好的地区, 其他物种对降水要求不高。最常见的 3 种有刺灌木(酸枣、小叶鼠李和红花锦鸡儿)都分布在相对温暖的地区,蔷薇属有刺灌木则主要分布在年均温低于 5 °C 的地区(图 4)。有实验研究证明多种蔷薇属植物具有良好的抗寒特性, 经历低于-45°C的低温仍可存活生长[25]。
酸枣是在京津冀山地分布最广泛的一种有刺灌木, 研究区内存在有刺灌木的样方中, 一半以上都有酸枣出现。酸枣为枣的变种, 属于泛热带成分的枣属, 主要起源于古南大陆, 但其分布范围延伸到温带地区, 表明了我国温带区系与热带的亲缘关 系[17,22]。酸枣具有深广的根系, 能够抗旱, 耐贫瘠, 在我国华北、西北、东北和华东的向阳山坡、丘陵和平原均有分布, 并因其水土保持的功能而作为绿化物种广泛栽培[23,26]。从其分布格局和与环境因子的关系可以发现, 酸枣主要分布在海拔较低、坡度较缓的阳坡和半阴坡, 其株数则表现为由阳坡至阴坡显著减少的特征, 也有研究证明酸枣在阳坡的生物量和根密度均明显大于阴坡, 荆条-酸枣灌丛的分布受坡向影响明显[26,27], 说明光照是限制酸枣分布的重要因子。有酸枣出现的样方集中在年降水量550 mm 左右, 年均温 10°C 左右, 降水和温度均与酸枣的植株高度有显著正相关关系, 而株数则与年均温呈现出显著负相关关系, 说明缺水会限制酸枣生长高度, 高温则会减少酸枣在群落中出现的数量。针对酸枣幼苗生长的实验研究表明, 土壤干旱胁迫对其侧枝生长、苗高和基径都有一定程度的限制[28]。但是, 酸枣的抗旱性较强, 能够通过降低氧化损伤、积累渗透调节物质和维持较高光合速率等途径来抵抗干旱[12]。
鼠李属是京津冀山地分布的有刺灌木种类最多的一个属, 包括7个物种, 出现在超过30%的有刺灌木样方内, 以小叶鼠李分布最为广泛。鼠李属植物的小枝顶端通常形成枝刺, 在旱生条件下往往表现出分枝增多, 枝刺增多、增大而坚硬的特性[5,23]。鼠李属为世界广布成分, 是灌丛的主要组成[17,22]。研究区内鼠李属有刺灌木的海拔分布范围较广, 在100~1400 m的山地均有出现, 主要分布在40°以内的阳坡、半阴坡和阴坡, 圆叶鼠李可生长在林下, 小叶鼠李也有一定的耐荫能力[23,29], 因此相对于酸枣而言, 出现在较多阴坡样方中。其株高表现出与年均温的显著负相关关系, 说明高温环境会抑制鼠李属有刺灌木的高生长。地形和气候因子与鼠李属有刺灌木的分布相关关系较小且多不显著, 可能与土壤条件有关。
研究区内的锦鸡儿属有刺灌木包括4种, 出现在122个样方中, 恒有度较高。锦鸡儿属为温带亚洲成分, 分布主要限于亚洲温带, 起源于古北大陆, 但发展历史并不古老, 其分化中心与亚洲干旱中心相吻合, 但旱化和特化的程度比中亚成分低, 在喜马拉雅山区和我国东部森林都有出现[17,22]。研究区内锦鸡儿属有刺灌木主要分布在海拔700 m以内的低山区阳坡和半阳坡, 仅树锦鸡儿一种在高海拔样方内曾出现, 不同种之间坡度分布差别较大。出现锦鸡儿属有刺灌木的样方年降水范围在 500~650 mm 之间, 年均温多高于 8°C, 气候相对暖湿。根据以往对锦鸡儿属抗旱性的研究, 红花锦鸡儿、树锦鸡儿和小叶锦鸡儿均属于喜暖的中生灌木类 型[30,31]。与主要分布在我国草原区和荒漠区的锦鸡儿属旱生、强旱生类型相比, 中生种叶面积较大, 叶片水势较高, 水分调节能力较弱, 主要分布在半湿润区, 并可凭借其活跃的代谢水平和快速生长而保持竞争优势[30]。本文结果表明其株数、盖度都与坡向、年降水呈显著正相关关系, 盖度与年均温呈显著负相关, 说明强光照、缺水和高温构成的干旱会抑制锦鸡儿属中生类型在群落中出现。
通过分析在京津冀山地选取的 460 个样地调查结果, 我们发现, 有刺灌木在研究区植被中较为常见, 出现在 60%的样地内, 并主要出现在荆条灌丛群落中, 在少数样地成为灌木层优势种。全部 105种灌木中有 24 种有刺种类, 主要来自鼠李科、蔷薇科和豆科。最常见的 3 种有刺灌木(酸枣、小叶鼠李、红花锦鸡儿)主要分布于海拔 600 m 以下的低山丘陵区, 常见于坡度较缓的阳坡、半阳坡, 及年均温高于 8°C 的温暖地区。最常见的 3 个属中, 光照和干旱是限制枣属、锦鸡儿属生长的重要因子, 鼠李属有刺灌木分布范围广, 受地形、气候因子限制较小。本文结果表明: 受气候、地形和群落因素的共同影响, 京津冀山地有刺灌木分布在多样化的生境条件之下, 但其中最常见的物种则主要出现在干燥的低山丘陵阳坡和半阳坡。
The authors have declared that no competing interests exist.
Changes in traits of shrub canopies across an aridity gradient in northern Patagonia, Argentina Are semiarid shrubs resilient to drought and grazing? Differences and similarities among species and habitats in a long-term study Resource pulses, species interactions, and diversity maintenance in arid and semi-arid environments 灌木群落及生物量研究综述 中国东北鼠李属植物形态、分类与地理分布 Flores G J A, Hernandez J, et al. The structure and function of vegetal ecosystems of semiarid areas in northeastern Mexico // Camilleri M L. Structural analysis of native plants: chapter 2
Lauenroth W K et al. Resource partitioning between shrubs and grasses in the Patagonian steppe From protégé to nurse plant: establishment of thorny shrubs in grazed temperate woodlands
Spiny plants in the archaeolo-gical record of Israel
酸枣生理生化特性对干旱胁迫的响应 北京八达岭地区灌木群落空间分布格局与土壤环境因子的关系分析 灌丛分布与坡度的关系: 京津冀山地实例研究 Very high resolution interpolated climate surfaces for global land areas 几种蔷薇属植物抗寒性指标的测定 酸枣的护坡性能及种子萌发研究[D] 山西太行山南段峡谷区荆条灌丛数量分析 土壤干旱对 2 个种源野生酸枣幼苗生长和生理特性的影响 内蒙古高原锦鸡儿属植物的形态和生理生态适应性 13 种锦鸡儿属植物叶的解剖生态学研究
/
1 研究区与研究方法
1.1 研究区概况
1.2 研究方法
科(共6科) 属(共10属) 种(代码) (共24种) 拉丁文名称 鼠李科 鼠李属 小叶鼠李(XYSL) Rhamnus parvifolia 圆叶鼠李(YYSL) Rhamnus globosa 东北鼠李(DBSL) Rhamnus schneideri var. manshurica 冻绿(DL) Rhamnus utilis 金刚鼠李* Rhamnus diamantiaca 锐齿鼠李* Rhamnus arguta 乌苏里鼠李* Rhamnus ussuriensis 枣属 酸枣(SZ) Ziziphus jujuba 豆科 锦鸡儿属 红花锦鸡儿(HHJJ) Caragana rosea 锦鸡儿(JJ) Caragana sinica 小叶锦鸡儿(XYJJ) Caragana microphylla 树锦鸡儿(SJJ) Caragana arborescens 槐属 白刺花* Sophora davidii 蔷薇科 蔷薇属 美蔷薇(MQW) Rosa bella 山刺玫(SCM) Rosa davurica 黄刺玫* Rosa xanthina 山蔷薇* Rosa sambucina 野蔷薇* Rosa multiflora 悬钩子属 牛叠肚(NDD) Rubus crataegifolius 五加科 五加属 刺五加(CWJ) Acanthopanax senticosus 楤木属 楤木* Aralia chinensis 小檗科 小檗属 掌刺小檗(ZCXB) Berberis koreana 黄芦木* Berberis amurensis 胡颓子科 沙棘属 沙棘(SJ) Hippophae rhamnoides
2 结果
2.1 有刺灌木各物种的科属信息及其在群落中的重要性
属 灌木名称 样方个数 株高/m 总株数 盖度/% 鼠李属 小叶鼠李 183 0.97 2212 3.75 圆叶鼠李 17 1.48 382 5.75 东北鼠李 7 1.53 85 5.43 冻绿 5 1.24 36 1.40 小计 209 1.03 2715 3.90 枣属 酸枣 362 1.24 5493 5.70 锦鸡儿属 红花锦鸡儿 85 0.94 1115 2.75 锦鸡儿 4 1.30 13 2.75 小叶锦鸡儿 10 1.10 145 3.00 树锦鸡儿 11 1.12 225 24.73 小计 122 0.98 1498 4.97 蔷薇属 美蔷薇 36 0.88 410 3.01 山刺玫 32 1.32 675 10.30 小计 67 1.09 1085 6.49 悬钩子属 牛叠肚 19 0.78 302 6.71 五加属 刺五加 10 1.22 135 4.61 小檗属 掌刺小檗 8 0.50 82 0.71 沙棘属 沙棘 4 0.98 20 8.63 总计 640 1.12 11330 5.16
2.2 有刺灌木各物种的地理环境分布
种类 样方个数 海拔/m 坡度/(°) 坡向/(°) 年降水/mm 年均温/°C 范围 均值 范围 均值 范围 均值 有刺灌木 640 87~2172 557.3 27.0 215.5 411~692 541.4 -2.7~12.2 8.7 全部灌木 1462 87~2172 853.6 39.6 157.3 410~701 538.3 -2.7~12.5 7.1
2.3 环境因子对有刺灌木分布的影响
种类 重要性指标 坡度 坡向 年降水 年均温 全部有刺灌木 株高 -0.010 0.076* 0.145*** 0.099** 株数 0.049 0.033 0.090* 0.018 盖度 -0.026 0.098** 0.116** 0.073* 枣属 株高 0.004 0.062 0.162** 0.129* 株数 0.046 -0.142** 0.052 -0.125* 盖度 -0.021 0.011 0.039 0.085 鼠李属 株高 -0.088 0.009 0.123 -0.143* 株数 0.095 0.074 0.084 0.019 盖度 0.066 -0.017 0.024 -0.030 锦鸡儿属 株高 0.032 0.158 -0.028 0.128 株数 0.082 0.281** 0.201* -0.102 盖度 0.001 0.211* 0.615*** -0.354***
3 讨论
3.1 京津冀山地有刺灌木分布格局
3.2 有刺灌木常见种属分布的影响因子
4 结论
参考文献
[1]
[2]
[3]
[4]
[5]
[6]
[7]
[8]
[9]
[10]
[11]
[12]
[13]
[14]
[15]
[16]
[17]
[18]
[19]
[20]
[21]
[22]
[23]
[24]
[25]
[26]
[27]
[28]
[29]
[30]
[31]

〈
〉
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}