
Ying LI
通讯作者:
收稿日期:
2016-02-11
修回日期:
2016-04-15
网络出版日期:
2017-05-11
版权声明:
2017 《北京大学学报(自然科学版)》编辑部 《北京大学学报(自然科学版)》编辑部 所有
基金资助:
展开
摘要 基于海北站氮磷添加实验平台, 研究青藏高原高寒草地常见植物叶片干物质含量(LDMC)、比叶面积(SLA)和叶片碳、氮、磷含量(LC, LN, LP)属性对N添加、P添加和NP同时添加的响应。从10个物种总体来看, N添加使LN显著增加9.4%。P添加使LP显著增加81.8%。N添加和P添加对LDMC和SLA有显著的交互作用。在无N添加条件下, P添加使LDMC减小2.3%, SLA增加3.5%; 在N添加条件下, P添加使LDMC减小10.1%, SLA增加15.3%。N添加和P添加对基于叶经济谱的物种排序无显著影响, 而NP同时添加显著改变了基于叶经济谱的物种排序。结果表明, 通过提高SLA和叶片NP含量, 使高寒草地植物对光、土壤可利用性N和P的获取与利用能力增强。NP同时添加可以显著改变叶属性的种间变异, 进而导致青藏高原高寒草地常见植物在叶经济谱上形成新的分布格局。因此, 在利用叶属性预测群落结构和生态系统功能对NP同时添加的响应时, 应该考虑物种的特异性响应。
关键词:
Abstract To investigate species’ resource competition strategies with traits of the leaf economics spectrum across contrasting environments and to examine the effects of nutrient additions on the ranking of species based on their leaf economics spectrum in an alpine grassland on the Qinghai-Tibetan Plateau, five leaf traits (LDMC: leaf dry matter content, SLA: specific leaf area, LC: leaf carbon concentration, LN: leaf nitrogen concentration and LP: leaf phosphorus concentration) were measured for 10 plant common species in all plots. The results showed that N addition significantly increased LN by 9.4% and P addition significantly increased LP by 81.8%. There were significant interactions between N addition and P addition on SLA and LDMC, which increased SLA by 15.3% and decreased LDMC by 10.1%. In addition, there were species-specific responses of leaf traits across multivariate trait space to nutrient additions. The variation in species responses to NP addition significantly changed the species ranking based on the leaf economics spectrum. These results showed that co-occurring species followed a conservative strategy in the infertile environment and an exploitative strategy in the fertilized ones by increasing SLA and leaf nitrogen and phosphorus concentrations. Different species responses to NP addition caused a new species distribution based on the leaf economics spectrum. These results suggest that, before using leaf traits to predict responses of community structure and ecosystem functioning to nitrogen and phosphorus additions, it is necessary to take the species-specific responses into consideration.
Keywords:
植物功能属性(plant functional traits)指间接地通过影响物种的“生长-繁殖-生存”三方面的个体表现,影响物种适合度的任何一种形态-生理-物候(morpho-physio-phenological)特征, 如植株高度、叶片大小、种子质量及单株生物量等[1]。这些功能属性相互关联且权衡, 进而通过影响植物特定的表现属性(performance traits), 如植物生物量和植物存活等, 最终影响个体适合度[1,2]。
植物功能属性能够反映植物在形态结构和生理特征等方面对环境变化的响应及适应策略。其中, 叶属性既能反映植物对资源的获取、利用和保存策略, 也能体现植物的生长对策[3], 同时, 叶属性可塑性大, 对环境变化的响应更为敏感而迅速, 易于测定, 便于将植物种群或群落所具有的相似格局扩展到区域或全球尺度上[4]。因此, 针对植物功能属性开展的研究主要集中在叶属性方面[5,6]。许多基于大尺度的研究发现, 叶属性间的多元关系中存在一个可以量化不同物种经济策略的主轴[5,7-9], 即在任何一个资源梯度下, 叶寿命长、资源利用成本高、氮含量低和光合速率低的物种生长缓慢, 叶寿命短、成本低、氮含量高和光合速率高的物种生长快速[5]。与植物经济策略相关的叶属性间协同变化、相互权衡的内在关系被称为叶经济谱(leaf economics spectrum) [5-6,10]。叶经济谱理论基于植物叶片的结构、化学和生理属性组合, 对物种进行排序, 从而量化不同物种的资源权衡策略。例如叶经济谱中基于促进植物快速生长的属性(如高的叶片氮、磷含量和比叶面积)与延长叶寿命以保存资源的属性(如高的叶片干物质含量)之间权衡的物种划分[5,10-11]。在全球变化背景下, 通过不同植被类型及其典型物种的叶属性的响应来研究和揭示生态系统功能的响应, 是当前植物生态学研究的热点问题 之一。
关于植物叶属性及其相互关系对土壤养分可利用性资源变化的响应, 已有许多研究[3,12-15], 但在不同研究尺度下得到的结果并不一致。在植被水平乃至全球范围的大尺度下, 土壤养分含量差异对基于叶属性的物种排序[16,17]、叶属性间异速生长和权衡关系的影响往往不大[5-6,18-19]; 在区域、群落或种群等小尺度下, 叶属性间的关系却依赖于土壤养分变化或群落类型[16,17]。He等[6]发现全球尺度下叶属性间的权衡关系同样存在于青藏高原高寒草地生态系统。在青藏高原高寒草地, 土壤可利用性氮(N)、磷(P)含量是植被生长的限制因子[16,17], 同时, 高寒草地植物叶片结构、化学计量学特征以及地上、地下功能关系具有一定的特殊性[16,17], 因此养分添加可能通过改变土壤养分可利用性而对不同物种的叶属性产生重要影响, 最终影响物种对变化环境的适应策略及生态系统功能。因此, 小尺度的施肥实验有助于深入揭示大尺度下物种叶属性对土壤养分变化的响应机理。
本研究以青藏高原高寒草地生态系统为研究对象, 通过野外养分调控(N添加、P 添加、NP 添加)实验平台, 探讨高寒草地常见植物叶片干物质含量(LDMC)、比叶面积(SLA)和叶片碳、氮、磷含量(LC, LN, LP)属性对N, P添加的响应及适应策略。如果N, P添加使LDMC和LC减小, 使叶片NP含量和 SLA 增加, 并且基于这 5 个属性的多元属性的第一主轴增加, 则表明高寒草地植物对光和土壤可利用性 NP 的获取与利用能力增强, 对资源保存能力减弱。如果不同物种基于叶经济谱的排序不发生改变, 则表明叶属性的种间变异不影响物种在叶经济谱上的分布格局。如果不同物种基于叶经济谱的排序发生显著变化, 则表明N, P添加可以显著改变叶属性的种间变异, 进而导致高寒草地植物在叶经济谱上形成新的分布格局。为了验证以上假说, 本研究拟解决两个问题: 1) N, P 添加后, 物种的资源竞争策略会由资源保存策略向资源快速获取与利用策略调整吗? 2) 多元叶属性对N, P添加的响应是否影响物种在叶经济谱上的分布格局?
本研究在青海海北高寒草地生态系统国家野外科学观测研究站(简称海北站)(37°29′—37°45′N, 101°12′—101°23′E)进行。该站地处青藏高原东北隅的青海海北藏族自治州门源县境内, 在祁连山北支冷龙岭东段南麓坡地的大通河河谷西段, 海拔3200~3600 m。站区属于典型的高原大陆性气候, 多年平均气温为-1.6°C, 年均降水量约 560 mm, 暖季(5—9月)短暂而凉爽湿润, 冷季(10月—翌年4月)漫长而寒冷干燥[28]。本研究选择长期适应于高寒环境下的高寒矮嵩草草甸为研究对象, 其植物群落以矮嵩草(Kobresia humilis)、异针茅(Stipa aliena)、垂穗披碱草(Elymus nutans)、麻花艽(Gentiana straminea)、美丽风毛菊(Saussurea pulchra)和高山豆(Tibetia himalaica)等为优势物种。土壤类型为高寒草甸土, 土壤中 N, P 总量丰富而可利用养分贫乏, 大多以有机态为主, 矿化作用较弱[29]。
本研究基于 2011 年在海北站建成的养分调控实验平台, 实验设计为完全随机区组设计, 共 4 种处理, 分别为对照、N 添加(10 g N/(m2·a))、P 添加 (5 g P/(m2·a))和 NP 同时添加(10 g N/(m2·a)+5 g P/ (m2·a))。所施氮肥为尿素, 磷肥为重过磷酸钙。每个处理设置6个重复, 共计24个样方, 样方面积为6 m × 6 m。区组间有宽 2 m 的缓冲区隔开, 样方间有宽 1 m 的缓冲区用来消除相邻处理的干扰。随机选取其中的4个重复开展实验。
实验期间, 将预先称好的肥料平均分成3份分别于2011年和2012年6月1日、7月1日和8月1日的傍晚均匀地撒在样方中。选择在傍晚施肥的目的是可以利用雨水或次日形成的露水, 使肥料迅速溶解进入土壤。对于每个实验小区, 分别选择群落内共有的 10 种常见植物为研究对象(表 1), 并将其分为禾类草和杂类草植物功能群。这 10 个常见种生物量之和占群落总地上生物量的75%以上。
表1 海北高寒草地10种常见植物基本特征
Table 1 Common species and their functional groups in the alpine meadow of Haibei, Qinghai, China
2012年8月初, 在所有实验小区内各物种选择3~5 株长势一致、健康完整的植株, 每个植株采集5~10 片成熟完整的叶片置于装有已吸水的滤纸的塑料袋中, 用冰盒冷藏带回实验室。叶片吸水饱和后取出, 并用吸水纸将叶片表面的水分吸干, 测定叶片鲜重(Lw)。然后用扫描仪(CanoScan LiDE 25)扫描叶片, 用 Image J软件(v2.1.4.7)测定其叶面积(La)。然后将叶片放置在 60°C 的烘箱中烘干至恒重(48 小时), 测定叶片干重(Ld)。将烘干的叶片样品研磨后, 一部分用 C/N 元素分析仪(2400II CHNS/O Elemental Analyzer, Perkin-Elmer, USA)测定叶片全碳、全氮含量(C%, N%), 其余的样品用浓H2SO4, H2O2和HF消煮后, 用钼蓝比色法测定叶片全磷含量(P%)[30]。
叶干物质含量(LDMC)、比叶面积(SLA)和单位质量的叶碳、氮、磷含量(LC, LN, LP) 等叶属性的计算公式如下:
LDMC (g/kg) = Ld / Lw,
SLA (cm2/g) = La / Ld,
LC (g/kg) = C%×单位换算系数,
LN (g/kg) = N%×单位换算系数,
LP (g/kg) = P%×单位换算系数,
式中, Ld和Lw分别为用于叶干物质含量测定的叶片干重和鲜重(g), La为通过Image J软件测定的叶面积(cm2)。
使用R3.1.1软件对上述数据进行统计分析。其中, 采用三因素方差分析(three-way ANOVA)检验 N 添加、P 添加和物种及其交互作用对于叶属性的影响; 采用主成分分析法(primary component analysis, PCA)和t检验分析物种的多元叶属性对N, P添加响应的一致性; 采用t检验分析N, P添加与对照处理下基于叶经济谱的物种排序的一致性。[PC1N addition-PC1Control, PC2N addition-PC2Control]表示物种在多元属性空间中对N添加的响应变化; [PC1P addition-PC1Control, PC2P addition-PC2Control]表示物种在多元属性空间中对P添加的响应变化; [PC1NP addition-PC1Control, PC2NP addition-PC2Control]表示物种在多元属性空间中对 NP 添加的响应变化[20, 31]。使用SigmaPlot10.0和R3.1.1软件作图。
表2 以N添加、P添加和物种为固定因子对叶干物质含量、比叶面积和叶片C, N, P含量的三因素方差分析
Table 2 Three-way ANOVA test of effects of N addition, P addition, species and their interactions on leaf dry matter content, specific leaf area, leaf C, N and P concentrations
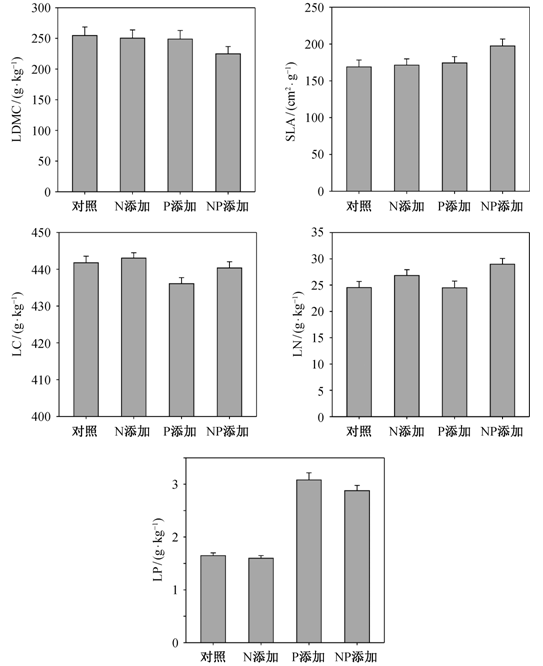
方差分析结果表明, 除LP外, N添加对所有叶属性均有显著影响(p<0.05; 表2)。除LN外, P 添加对所有叶属性均有显著影响(p<0.05; 表2)。N添加主要使 LN 增加 9.4% (图 1; 表 3), 其中禾草类 增加12.5%, 杂类草增加8.3% (表4); P添加主要使LP增加81.8%(图 1; 表 3), 其中禾草类增加103% (p<0.001), 杂类草增加74.9% (p<0.001)(表4)。N添加和P添加对LDMC和SLA有显著的交互作用(p<0.05; 表2)。在不添加N的条件下, P添加使LDMC 减小 2.3%, SLA 增加 3.5% (图 1; 表 3); 在 N 添加的条件下, P 添加使 LDMC 减小 10.1%, SLA增加15.3%(图 1; 表4)。
图1 海北高寒草地常见种的LDMC, SLA, LC, LN和LP对N添加、P添加和NP添加的响应(平均值±标准误差) Fig. 1 Responses of LDMC, SLA, LC, LN and LP to nitrogen and phosphorus additions for common species (mean±SE)
表3 海北高寒草地常见植物的LDMC, SLA, LC, LN和LP对N添加、P添加和NP添加的响应(平均值±标准误差)
Table 3 Responses of LDMC, SLA, LC, LN and LP to nutrient additions for common species (mean±SE)
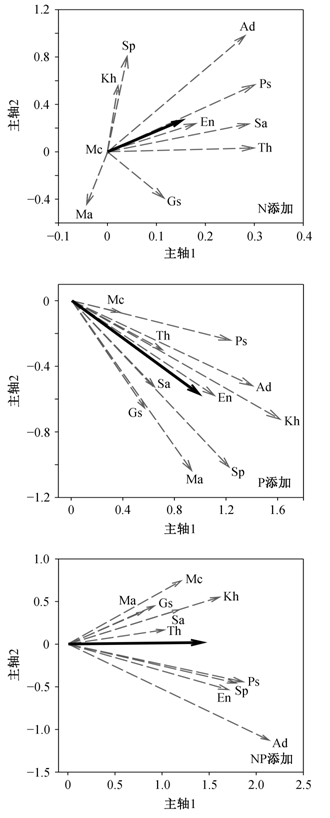
主成分分析结果表明(图 2), 主成分 1 和 2 解释的方差分别为总方差的 47.9%和 26.6%, 合计74.5%; 即主成分 1 和 2 可解释原始 5 个变量的74.5%的变异。其中主成分 1 与 SLA 和 LP 的相关性较高,主成分 2 主要与 LC 相关性较高。LDMC和LN则既与主成分1相关, 又与主成分2相关。
图2 不同N、P添加条件下海北高寒草地常见种在多元叶属性空间内的排序(Sa: 异针茅; En: 垂穗披碱草; Kh: 矮嵩草; Ma: 青海苜蓿; Th: 高山豆; Gs: 麻花艽; Sp: 美丽风毛菊; Mc: 圆萼刺参; Ad: 重冠紫菀; Ps: 钉柱委陵菜) Fig. 2 Common species ranking based on traits of the leaf economic spectrum under different nitrogen and phosphorus conditions in the alpine grassland in Haibei, Qinghai, China
表4 海北高寒草地禾草类植物和杂类草植物的LDMC, SLA, LC, LN和LP对N添加、P添加和NP添加的响应(平均值±标准误差)
Table 4 Responses of LDMC, SLA, LC, LN and LP to nutrient additions for graminoids and forbs (mean ±SE)
由物种在多元属性空间内的响应(图 3) 可知, 高寒草地常见植物在叶属性对 N, P 添加的响应上具有物种特异性。在 N 添加处理下, 土壤中 N 的可利用性从低到高的变化使叶属性在物种间呈现出较大变异, 从而导致叶属性均值在 PCA 主轴 1 和主轴 2 上均无显著变化(图 3(a)), 因此, 基于 PCA主轴 1 的物种排序无明显变化(p>0.05, t检验)(表5)。在 P 添加处理下, P 的可利用性从低到高的变化使物种一致性地沿主轴1的正向和主轴2的负向改变叶属性, 但叶属性均值在 PCA 主轴 1 和主轴 2上的变化均不显著(图 3(b)), 即基于主轴 1 的物种排序的变化不显著(p>0.05, t检验)(表 5)。在 NP 添加处理下, N 和 P 从低的可利用性到高的可利用性的变化使叶属性均值在主轴 1 上显著增加, 也就是基于主轴 1 的物种排序显著变化(p<0.05, t 检验) (表 5), 但物种在主轴 2 上分成两个类群沿相反方向响应, 进而导致叶属性均值在主轴 2 上变化不显著(图3(c))。
表5 海北高寒草地10种常见植物在4种养分添加处理下基于主成分分析主轴1的排序
Table 5 Rankings of ten common species based on the PCA axis 1 in the four nutrient additions treatments in the alpine meadow of Haibei, Qinghai
由10个物种基于前两个主因子得分的排序可知(图 2; 表 5), 在对照处理下, 第一因子得分最高的是豆科杂类草高山豆(Th)和青海苜蓿(Ma), 得分最低的是禾草类异针茅(Sa)、矮嵩草(Kh)和垂穗披碱草(En); 第二因子得分最高的是非豆科杂类草圆萼刺参(Mc)、美丽风毛菊(Sp)和重冠紫菀(Ad), 得分最低的是青海苜蓿(Ma), 表明高山豆和青海苜蓿相比于异针茅、矮嵩草和垂穗披碱草, 叶片更薄, 光合能力更强, 因此更倾向于快速获取和利用资源的竞争策略。在N添加和P添加处理下, 10 个物种基于叶经济谱(主轴1)的排序不发生显著变化, 表明多元叶属性对 N 添加和 P 添加的响应不影响物种在叶经济策略上的差异。与此相反, 在 NP 添加处理下, 多元叶属性的响应使物种基于叶经济谱的排序发生显著变化, 第一因子得分最高的是高山豆(Th)和重冠紫菀(Ad), 得分最低的是异针茅(Sa)、矮嵩草(Kh)和麻花艽(Gs), 表明 NP 添加可以显著改变叶经济策略相关属性的种间变异。在土壤可利用性N, P增加条件下, 高山豆和重冠紫菀成为资源快速获取与利用能力最强的物种, 异针茅、矮嵩草和麻花艽成为资源保存能力最强的物种。
图3 海北高寒草地常见种在多元叶属性空间内对 N 添加、P添加和NP添加的响应(灰色矢量代表 N, P 添加后各物种在属性空间中的位置变化, 黑色矢量代表N, P添加后物种的平均位置变化。物种缩写同 Fig. 3 Shifts of species leaf traits across multivariate trait space in response to N addition, P addition and NP addition in the alpine grassland in Haibei, Qinghai
结果表明, 在 N 或 P 添加的群落内, 物种具有较高的 SLA, LN 和 LP, 而在对照群落内, 物种具有较高的 LDMC 和LC。LN 和 LP 对 N, P 添加的正响应与已有的研究[32,33,34,35,36,37]结果一致。但是, SLA 和LDMC 对 N, P 添加的响应具有不确定性。万宏伟等[14]发现, N 添加可以增加内蒙古草原植物的SLA。赵新风等[38]发现, N 添加使荒漠草原植物钠猪毛菜SLA减小, LDMC 增加。然而, 本研究发现 NP 添加使高寒草地植物 SLA 增加, LDMC 减小。同时, NP 添加显著增加了多元叶属性主轴 1 均值, 对主轴 2 均值无显著影响。以上结果表明, 高寒草地物种通过叶属性间的异速变化来响应土壤可利用N, P 资源的增加, 这与许多已有研究结果[18,22,34] 一致。
主成分分析结果表明, 主轴 1 主要反映 SLA, LN 和 LP。SLA 是反映植物碳收获策略的关键叶属性之一, 与植物潜在的相对生长速率、单位质量光合速率和单位质量叶 N 含量正相关[5,39-40]。LN和 LP 反映植物对土壤养分的吸收特性[7], 与单位质量光合速率呈正相关关系[5,6]。因此, 在某一土壤资源梯度下, SLA, LN 或 LP 较高的物种具有较强的N, P吸收和光捕获能力, 从而表现为快速生长策略[10], 以达到植物自身生存、生长和繁殖的目的。主轴 2 主要反映 LC 和 LDMC。LDMC 和 LC对植物养分保存能力具有很好的指示作用, 反映植物对养分贫瘠生境的适应策略, 且 LDMC 与 SLA具有显著的负相关性[16,41-42]。因此, LDMC 或 LC较高的植物具有较强的养分保持能力和对资源有限环境的适应能力[43], 从而表现为缓慢生长策略[10]。
从物种在叶经济谱上的位置来看, 物种在低的可利用性N, P 环境中采取资源保守策略, 在高的可利用性N, P 环境中则倾向于采取资源快速获取与利用策略。因此, 在低养分环境中具有较高 SLA和 LN 的快速生长物种(如高山豆), 在高养分环境中仍具有相对较高的 SLA 和 LN, 以维持快速生长能力; 在低营养环境中具有较高 LDMC 和 LC 的缓慢生长物种(如异针茅和矮嵩草), 在高养分环境中仍具有相对较高的 LDMC 和 LC, 以维持养分保持能力。但是多元叶属性对NP添加的响应显著改变了叶经济策略相关属性的种间变异, 进而导致物种在叶经济策略上的差异发生变化。与杂类草相比, 禾草植物快速获取与利用资源的能力较弱(图 2), 但我们发现垂穗披碱草(En)等禾草植物的叶片N, P含量对 NP 添加的响应比杂类草植物更明显(LN, 27.0% vs. 15.2%; LP, 88.9% vs. 70.0%), 进而导致禾草植物的资源快速获取与利用能力的提升更明显, 这与禾草植物的“奢侈消费”理论相一致, 可能是禾草植物为了限制其他快速生长的共存杂类草获取和利用可利用性N, P资源的竞争策略之一[44,45]。
从多元叶属性的响应来看, 土壤可利用N, P含量从低到高的变化使不同物种在多维属性空间内的响应幅度和响应方向呈现出差异性。本研究发现, 叶属性在对 N 添加的响应上变异最大。我们认为物种对 N 添加响应的趋异性可能是由于以下原因。1) 同一属性在不同物种的功能属性空间中所占的比重不同, 导致基于属性的生态位的多维度组成具有种间差异性。2) 当土壤可利用性 N 增加时, 可利用性P对植物生长的限制作用相对增强。群落内共存物种对可利用性 P 和光资源的竞争加强, 导致物种基于属性的生态位发生分化, 进而促进叶属性响应的种间变异以维持彼此对环境资源的获取、保存与利用。3) 叶属性的可塑性具有物种特异性, 且不同物种的叶属性对环境变化的敏感性也存在差异[46,47]。这与 Wright 等[20]通过控制实验研究湿地物种对N添加的响应结果相似。Adamidis 等[31]发现物种对生境变化的响应也具有广泛的差异, 导致叶经济谱无显著变化。然而, Lienin 等[15]发现土壤养分变化使得物种叶属性的响应具有趋同性并且对植物叶经济谱产生重要影响。本研究也发现物种多元叶属性对P添加和NP添加的响应具有趋同性。这可能是由于环境骤变后植物群落在重新构建过程中新的环境筛选导致共存物种的叶属性趋同[48]。同时, 叶属性的种内变异对物种响应环境变化的影响也不可忽视[49]。此外, 还有可能是因为我们所研究的属性对于区分物种适应环境变化的策略差异的能力较弱, 植物的根和茎属性可能对土壤可利用性养分和光资源的竞争更重要。叶属性对 N, P 添加响应的种间差异性与许多大尺度研究结果[5,50]不一致, 还有待在其他群落类型或功能型物种间进行验证和深入研究。
总体来看, 高寒草地常见植物的 SLA, LN 和LP随土壤可利用性N, P增加而增大, LDMC和LC随土壤可利用性N, P增加而减小, 多元叶属性也随土壤可利用性N, P增加而增大, 表明本地共存物种在资源竞争策略上正在由资源保守向资源快速获取与利用方向调整, 进而调节其生长速率和资源竞争力, 以适应当前的无机环境及种间竞争。N 添加和P 添加对高寒草地常见植物基于叶经济谱的排序无显著影响, 而 NP 添加显著改变了高寒草地常见植物基于叶经济谱的排序, 表明多元叶属性对 NP 添加的响应使叶经济策略相关属性的种间变异发生明显变化, 进而导致本地共存物种在叶经济谱上产生了新的分布格局。在土壤可利用性 N, P 受限的条件下, 资源快速获取与利用能力最强的是高山豆和米口袋; 资源保存能力最强的是异针茅、矮嵩草和垂穗披碱草。在土壤可利用性 N, P 充分的条件下, 高山豆和重冠紫菀在叶经济谱的“快速投资-收益”端, 成为资源快速获取与利用能力最强的物种; 异针茅、矮嵩草和麻花艽在叶经济谱的“缓慢投资-收益”端, 成为资源保存能力最强的物种。
我们的研究结果与长期 N, P 添加的实验结 果[51,52,53]相一致, 说明高寒草地生态系统经历长期的养分输入后, 杂类草植物在资源快速获取与利用方面可能不再具有明显的优势地位, 进而被更适应高养分环境的禾类草植物所替代。
致谢 感谢青海海北高寒草地生态系统国家野外科学观测研究站的支持。
The authors have declared that no competing interests exist.
Let the concept of trait be functional! Morphology, performance and fitness 植物功能性状与环境和生态系统功能 植物叶寿命及其相关叶性状的生态学研究进展 The worldwide leaf economics spectrum A test of the generality of leaf trait relationships on the Tibetan Plateau Hunt R et al. Integrated screening validates primary axes of specialisation in plants Generality of leaf trait relationships: a test across six biomes
The plant traits that drive ecosystems: evidence from three continents The world-wide ‘fast-slow’ plant econo-mics spectrum: a traits manifesto Taxonomic, phylogenetic, and environmental trade-offs between leaf productivity and persistence Evolutionary divergences in leaf structure and che-mistry, comparing rainfall and soilnutrient gradients Climate change in the Arctic: using plant functional types in a meta-analysis of field experiments
羊草草原群落 6 种植物叶片功能特性对氮素添加的响应 Consistency of species ranking based on functional leaf traits Variation in leaf traits through seasons and N-availability levels and its consequences for ranking grassland species A global study of relationships between leaf traits, climate and soil measures of nutrient fertility Biogeographic constraints on the world-wide leaf economics spectrum Does the leaf economic spectrum hold within local species pools across varying environmental conditions? Disentangling coordination among functional traits using an indivi-dual-centred model: impact on plant performance at intra-and inter-specific levels
Intraspecific growth and functional leaf trait responses to natural soil resource gradients for conifer species with contrasting leaf habit
青藏高原高寒草甸植物群落生物量对氮、磷添加的响应
青藏高原高寒草地和内蒙古高原温带草地主要双子叶植物叶片解剖特征的比较研究
Alpine climate alters the relationships between leaf and root morphological traits but not chemical traits
Stoichiometry of root and leaf nitrogen and phosphorus in a dry alpine steppe on the northern Tibetan Plateau
海北高寒草甸生态系统定位站气候、植被生产力背景的分析 青藏高原高寒草甸生态系统管理的生态学基础: 海北高寒草甸生态系统研究站 Methods of soil analysis, Part 3, Chemical methods Species adaptive strategies and leaf economic relationships across serpentine and non-serpentine habitats on Lesbos, eastern Mediterranean Differential responses to nitrogen fertilization in native shrubs and exotic annuals common to Mediterranean coastal sage scrub of California Nitrogen and phosphorus enhance growth and luxury consumption of four secondary forest tree species in Borneo Global response patterns of terrestrial plant species to nitrogen addition 基于物种性状的两种高寒草甸优势禾草对施肥的响应 氮肥添加对青藏高原高寒草甸6个群落优势种生态化学计量学特征的影响 氮磷硅添加对青藏高原高寒草甸垂穗披碱草叶片碳氮磷的影响 养分与水分添加对荒漠草地植物钠猪毛菜功能性状的影响 Carbon and nitrogen content of congeneric annual and perennial grass species: relationships with growth. Plant, Relationships between leaf lifespan and structural defences in a low-nutrient, sclerophyll flora Thompson K E N, Hodgson J G. Specific leaf area and leaf dry matter content as alternative predictors of plant strategies Specific leaf area and leaf dry matter content of plants growing in sand dunes Chapin III F S. The mineral nutrition of wild plants revisited: a re-evaluation of processes and patterns The mineral nutrition of wild plants Luxury consumption of soil nutrients: a possible competitive strategy in above-ground and below-ground biomass allocation and root morphology for slow-growing arctic vegetation? Leaf-and shoot-level plasticity in response to different nutrient and water availabilities The role of plant traits and their plasticity in the response of pasture grasses to nutrients and cutting frequency Shifts in trait-combinations along rainfall and phosphorus gradients Within-species correlations in leaf traits of three boreal plant species along a latitudinal gradient Mor-phological and physiological adjustment to N and P fertilization in nutrient-limited Metrosideros polymor-pha canopy trees in Hawaii Contrasting effects of long term versus short-term nitrogen addition on photosynthesis and respiration in the Arctic
/
1 材料和方法
1.1 研究地点概况
1.2 实验设计
物种 科 功能群 异针茅Stipa aliena 禾本科 多年生禾草类 垂穗披碱草Elymus nutans 禾本科 多年生禾草类 矮嵩草Kobresia humilis 莎草科 多年生禾草类 麻花艽Gentiana straminea 龙胆科 多年生杂类草 美丽风毛菊Saussurea pulchra 菊科 多年生杂类草 高山豆Tibetia himalaica 豆科 多年生杂类草 青海苜蓿Medicago archiducis-nicolai 豆科 多年生杂类草 圆萼刺参Morina chinensis 川续断科 多年生杂类草 钉柱委陵菜Potentilla saundersiana 蔷薇科 多年生杂类草 重冠紫菀Aster diplostephioides 菊科 多年生杂类草
1.3 叶干物质含量、比叶面积和叶片 C, N, P含量的测定方法
1.4 数据统计分析
变异来源 df LDMC SLA LC LN LP N添加 (N) 1 28.24*** 12.80** 12.00** 37.30*** 3.61 P添加 (P) 1 60.61*** 31.70*** 33.00*** 3.49 1081.42*** Species (S) 9 656.03*** 144.82*** 58.00*** 174.55*** 50.37*** N×P 1 18.85** 11.08** 3.00 4.13 0.04 N×S 9 1.25 1.33 4.00*** 12.46*** 0.87 P×S 9 0.98 1.34 1.00 2.17* 16.66*** N×P×S 9 0.63 0.66 1.00 2.13* 1.31
2 结果
2.1 氮、磷添加对叶属性的影响
处理 LDMC/(g · kg-1) SLA/ (cm2 · g-1) LC/(g · kg-1) LN/(g · kg-1) LP/(g · kg-1) 对照 254.66±13.94 168.94±9.30 441.77±1.78 24.53±1.18 1.65±0.05 N添加 250.21±13.54 171.29±8.52 443.03±1.46 26.83±1.09 1.60±0.05 P添加 248.83±13.99 174.41±8.44 436.08±1.65 24.48±1.28 3.08±0.13 NP添加 224.83±11.84 197.46±9.26 440.35±1.71 28.98±1.10 2.88±0.10
2.2 主成分分析
功能群 处理 LDMC/(g·kg-1) SLA/ (cm2·g-1) LC/(g·kg-1) LN/(g·kg-1) LP/(g·kg-1) 禾草类 对照 360.12±16.95 127.73±10.92 446.90±1.24a 20.03±0.62b 1.35±0.08b N添加 359.18±14.86 133.81±11.47 448.51±1.21a 22.54±0.61ab 1.29±0.06b P添加 358.96±17.93 137.50±12.10 440.48±1.58b 19.83±0.84b 2.74±0.17a NP添加 318.01±11.50 158.31±16.29 447.10±1.60a 25.42±1.04a 2.54±0.12a 杂类草 对照 207.97±8.42 187.97±10.58 439.57±2.35 26.46±1.50 1.78±0.04b N添加 203.51±7.81 187.35±9.55 440.69±1.83 28.67±1.39 1.73±0.05b P添加 201.63±7.92 190.23±9.33 434.19±2.14 26.48±1.63 3.11±0.14a NP添加 184.90±7.88 214.24±9.57 437.45±2.10 30.50±1.40 3.02±0.12a
2.3 多元叶属性对氮、磷添加的响应与基于叶经济谱的物种排序
处理 物种排序 1 2 3 4 5 6 7 8 9 10 对照 Sa Kh En Gs Mc Ps Sp Ad Ma Th N添加 Sa Kh En Gs Mc Ps Sp Ad Ma Th P添加 Sa Mc Gs Kh En Ps Sp Ad Ma Th NP添加 Sa Kh Gs Mc En Ps Sp Ma Ad Th
3 讨论
参考文献
[1]
[2]
[3]
[4]
[5]
[6]
[7]
[8]
[9]
[10]
[11]
[12]
[13]
[14]
[15]
[16]
[17]
[18]
[19]
[20]
[21]
[22]
[23]
[24]
[25]
[26]
[27]
[28]
[29]
[30]
[31]
[32]
[33]
[34]
[35]
[36]
[37]
[38]
[39]
[40]
[41]
[42]
[43]
[44]
[45]
[46]
[47]
[48]
[49]
[50]
[51]
[52]
[53]

〈
〉

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}