
李伟 , 毛海颖
, 毛海颖
Wei LI
中图分类号:
S758
通讯作者:
Corresponding authors:
收稿日期:
2015-08-25
修回日期:
2015-12-26
网络出版日期:
2016-04-12
版权声明:
2017 北京大学
基金资助:
展开
摘要 为了研究树木的生长是否也存在突变的问题, 将突变论的尖角模型理论应用于我国北方——内蒙古赤峰市旺业甸林场 10 个常见树种单木材积的测定和计算。选取 10 个北方常见树种, 对其树高(H)、胸径(D)、材积(V)、地径(D0)等参数进行测定, 并以此作为真值, 利用改进差分进化算法对建立的“材积-树高-胸径”模型进行内符合精度检验, 利用边缘树种对模型进行外符合精度检验, 比较模型计算值与仪器测量值之间的差异。在经过一定的迭代之后, 结果表明: 模型的总体相对误差 RS 在[0.001, 0.05]范围, 平均相对误差 E 在 [-0.11, 0.02]范围, 总体预估精度 P 大于 80%, 说明模型拟合较好, 即突变论中的尖角模型理论适用于我国北方 10 个树种单木材积模型的建立, 同时说明随着树木的不断生长, 当树高达到一定高度时, 材积将遵循突变论的尖角模型理论。这是突变论的尖角模型理论首次应用到单木材积的测定, 并得到较好的拟合结果。同时, 在不连续的测量状态下建立的突变低维模型为树木材积的研究和各树种间的空间竞争研究提供了一定的理论和实践依据。
关键词:
Abstract In order to study whether the growth of trees existence mutations problems, measurement and calculation of the mutation based on the cusp model theory is applied to 10 common species single tree volume in Wang Ye Dian forest farm which is located in Chifeng City, Inner Mongolia, China. 10 northern common tree species were selected to carry out the experiment, the tree height (H), DBH (D), volume (V), diameter (D0) were measured which as the true value. To test the Internal and external accord accuracy of the “V-H-D” model by using the improved differential evolution algorithm and edge species and then compare the difference between model calculation value and instrument measuring. After a certain iteration, the results showed that the overall relative error (RS) of the model is in the range of [0.001, 0.05], the average relative error (E) is in the range of [-0.11, 0.02], and the overall prediction accuracy (P) is greater than 80%. The outcome had shown the good fitness of the model, namely the cusp catastrophe theory model applies to northern China 10 single species tree volume. With the growing of trees, when the height of the tree reaches a certain height, the volume will follow the theory model of cusp catastrophe theory. This is the cusp catastrophe theory model was first applied to the determination of single tree volume and got a better fitting result. Meanwhile, a low-dimensional model of mutation was established in the discontinuous measurement, which provides a theoretical and practical basis for the study of tree volume and various species of spatial competition.
Keywords:
自然界中很多有趣的现象涉及不连续性, 这种不连续性包括时间和空间范畴[1]。生态系统变化的不连续性表现在时间上, 主要包括细胞的分裂和重组、病虫害的发生和预防、森林火灾的蔓延及控制等。在空间上, 则包括种内与种间的竞争和入侵、捕食与被捕食的关系、菌类的生长与控制等[2]。在过去几十年中, 科学家们利用连续性的微积分状态方程建立各种数学模型来分析和解释连续变化的曲线, 虽然取得一定的成果, 但在分析像生态系统这样不连续性的情况时, 总会忽略一些相关因素, 分析结果不能全面地解释各类现象[3-4], 因此需要能够解决不连续问题的数学方法。1972 年法国数学家Thom[5]提出突变理论, 它是拓扑学的一个分支, 主要研究动态系统中的不连续性现象。突变理论的提出引起世界性的轰动和关注, 开辟了生态学的新方向。
突变论是关于奇点的理论, 适用于以下两种情况: 一是对理论的严格应用, 另一种是只利用突变论的思想和方法[6]。1976年, Jones等[7]将突变理论应用于渔场的管理, 对南极鳍鲸群体的大小变化进行解释。1977年, Jones[8]应用突变理论研究东加拿大云杉卷叶蛾森林生态系统, 以构造折迭突变模型来解释云杉卷叶蛾突发现象, 并采取一定防治措施。Loehle[9]运用突变论中的蝴蝶突变模型研究牲畜-草场生态系统。李勇等[10]将尖点突变模型应用于油气储层的边界分析, 从而有效地确定油气储层异常体的空间分布范围及边界。
目前, 国内外对突变论的应用主要集中在生态环境脆弱性研究[11-12]。南太平洋应用地球科学委员会(SOPAC)近年来致力于建立全球性的生态环境脆弱性指数工程[13], 主要包括病虫害的发生模型研究、风险灾害的RH模型研究、压力释放模型PAR的研究、地方性灾害HOP模型的研究等。在国内, 王瑞燕等[14]利用熵、突变论等非线性科学理论并结合遥感技术研究我国黄河三角洲区域的生态环境脆弱性问题, 得出该地区生态环境的变化同时与时间和空间有关的结论。此外, 在森林火灾的蔓延及预防方面, 孙玉荣等[15]结合BP神经网络对林火蔓延趋势进行预测。对突变论模型的应用研究已经覆盖生物、能源、环境等多个领域。
森林生态系统是环境保护的重点, 但是, 对于森林生态系统的研究却很少。本文将突变理论引入森林树种的种间生长竞争, 力求在森林生态系统研究领域获得突破。同时, 由于本文涉及的树种较多, 数据量相对庞大, 数据处理相对复杂, 因此引入遗传算法。遗传算法具有内在的隐并行性和更好的全局寻优能力, 采用概率化的寻优方法, 能自动获取和指导优化的搜索空间, 自适应地调整搜索方向, 不需要确定的规则。遗传算法具有克服局部极值陷阱和模式欺骗、提高全局最优的能力, 但是, 由于个体的种类繁多和选择的压力导致过早的收敛, 而使精度降低。为了克服遗传算法的这一缺点, 研究者纷纷寻找其他算法与遗传算法相结合, 例如强明等[16]将 BP 神经元网络与遗传算法结合进行 GPS 高程拟合, 获得较高的精度。因此, 本研究将使用遗传算法中的改进差分进化算法进行因子的提取和模型的建立, 即进行算子的选择来克服遗传算法的缺点, 在缩短计算时间的同时, 能够精确地筛选相关因子。
自然林的生长普遍存在树种间的竞争, 这种竞争虽然不像病虫害入侵那样, 在较短时间内有十分明显的表现, 但随着树木的生长, 存在一种树种逐渐被另一树种取代的现象。这类现象的发生影响种群内优势种的变化, 进而影响整个区域内生态系统的变化, 乃至区域小环境的变化[17]。例如, 火炬树作为一种引入种, 近年来在北京的一些区域, 已经取代当地树种成为优势树种。林业学者对此展开一系列研究, 并采取野外调查等方法, 对其属于引入种还是入侵种进行探讨。
本文以内蒙古赤峰市旺业甸林场为例, 利用电子经纬仪、测树枪等仪器, 随机测量林场内 10 类树种、1000 棵树的基本生长指标, 并以突变理论为依据, 将尖角模型引入单木的生长过程, 同时引入改进差分进化算法参与模型的建立和精度检验。今后, 可通过观测单木生长材积与树冠表面积等参数来预测种群中的优势树种, 并针对这种趋势采取有效的措施进行森林保护。本研究可为森林生态系统的有效循环提供一定的理论依据。
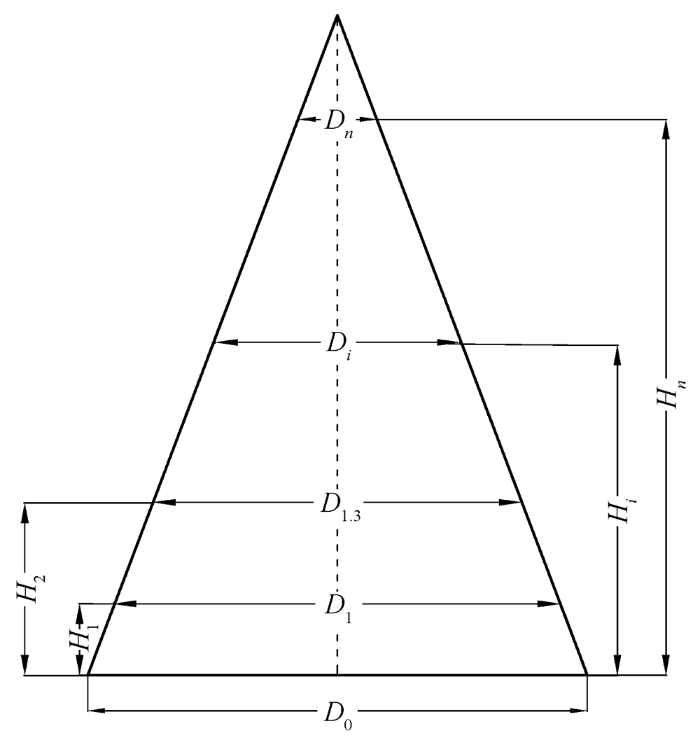
从树的底部开始测量, 树高H随着高度增加而增加, 即从H0增加到Hi (i=0, 1, 2, … n-1, n)。H0, H1, H2, H1.3, H3, … Hi分别代表从地面零点处到通过电子经纬仪测量的直到视线不能抵达处的高度, 其中, 胸径 1.3 m 处的树高为 H1.3。随着树高的增加, 胸径也随之增加, 胸径用 D 表示, 树高为H0的地面处的胸径(即所谓的地径) 用 D0 表示, 在整个测量过程中地径 D0 是不变的。树高1.3 m 处的胸径用D1.3表示。随着树高的增加, 胸径由D0逐渐增加到Dj (j=0, 1, 2, … n-1, n), 即D0, D1, D2, D1.3, D3, ... Dj。在这里, 树高H、胸径D和地径D0都是通过仪器测定直接获取的数据。随着树高的增加, 树形逐渐趋向三角形, 树高的顶点逐渐趋向三角形的顶点, 如图1所示。
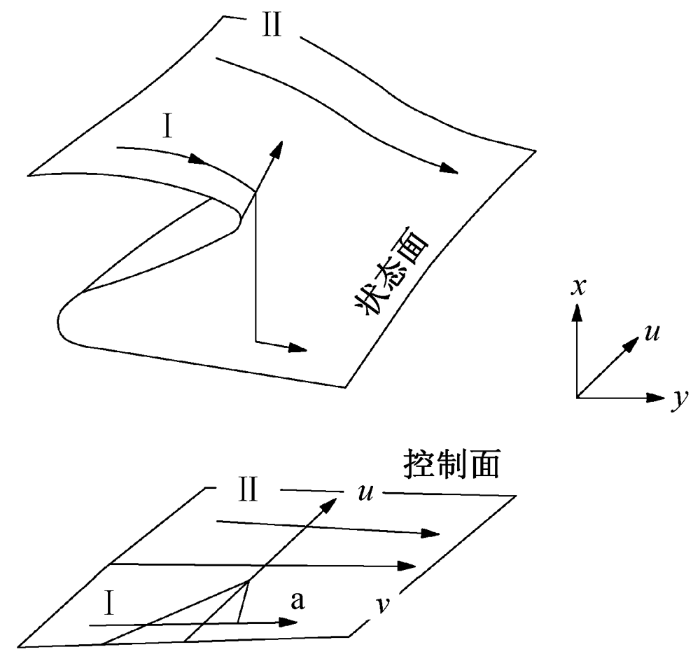
根据突变论中的尖角模型理论, 在假定的系统内, 假设任意时刻的状态都可以完全由给定的 n +1个变量(x0, x1, x2, x3, … xn)的值决定, n可以是有限的, 也可以是无限的。同时, 假定系统受到m+1个独立变量(P0, P1, P2, P3, … Pm)控制, 这些变量决定x的值, 即V(xi, P)。尖角模型空间及平面的变化轨迹如图2所示。
图2 尖角模型空间及平面的变化轨迹 Fig. 2 Sharp changes in model space and flat trajectory
本实验中, 树高 H、胸径 D 和地径 D0 都可看做是已知量(即控制变量), 要求的树木体积 V 是未知量(即状态变量), 主要由H, D和D0决定。在这里, 可以利用模型
V (x)=x4+Dx2+D0x , (1)
将单株树木的体积 V 的模型看成一个地径 D0和胸径D关于树高Hi (i=0, 1, 2, … n)的二元四次方程(式中 x 为树高 Hi), 通过将不同高度的树高、胸径和地径代入模型中, 得到不同高度处不同树种树木的材积V。
V° (x)= 0时, 4x3+2Dx+D0=0 ; (2)
V″(x)= 0时, 12x2+2D0=0。 (3)
式(1)是尖角模型的势函数, 式(2)是式(1)在平面中的投影, 即平衡曲面。式(2)与(3)联立, 可消去状态变量x。
本实验利用突变论的原理和尖角突变的模型来分析在树高H、地径D0和胸径D可测的情况下, 各类树种的体积变化情况, 对比分析树种间可能存在的种间竞争关系以及在未来可能存在的优势树种被取代的现象, 从而预测整个生态系统未来的发展动态及具体走向。
遗传算法(genetic algorithm)是模拟达尔文生物进化论的自然选择和遗传学机理的生物进化过程的计算模型, 是一种通过模拟自然进化过程获取最优解的方法, 其优势在于可避免局部最优解, 从而提取重要影响因子。遗传算法的基本模型为
$\max f(x)\ \ \text{s}\text{.t}\text{.}\ X\in R,\ R\in U$
其中, x=[x1, x2, …, xn]T为决策变量, U为基本空间, s.t. x为约束条件, R表示所有满足约束条件的解的集合, f (x)为目标函数[18]。
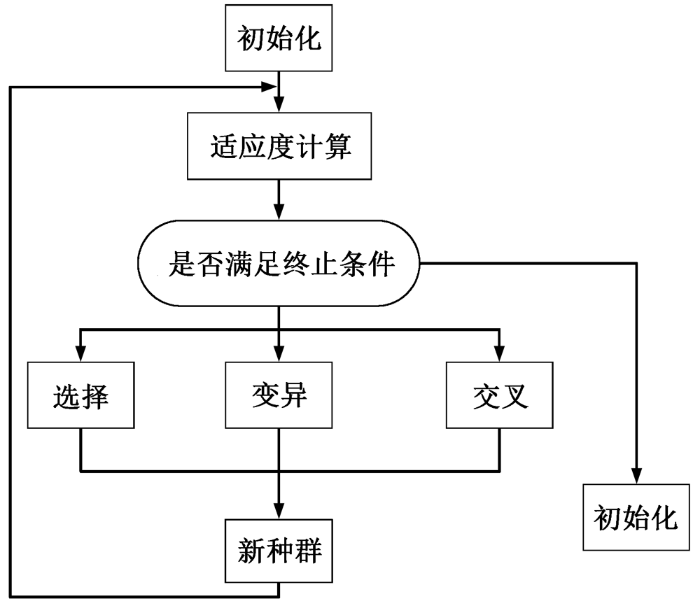
为了求得满足条件的最优解, 基础遗传算法采用人工进化、全局并行搜索的方式对问题进行优化搜索, 标准遗传算法的流程见图3。
进行计算之前, 首先确定对优化参数的编码方案。产生初始种群之后, 计算适应度值。适应度是判别个体优劣的标准, 为非负, 值越大表示优化越好。经过最优的迭代次数之后, 经过一系列的选择、交叉和变异, 选择个体形成交配池, 并对这些个体进行遗传运算而产生子代新种群。很多研究者利用遗传算法在种群个体的“优胜劣汰”问题上取得一定成效[19]。种群的多样性和选择压力是导致遗传算法过早收敛, 达不到最优收敛的主要因素。为了解决这两个弊端, 差分进化算法应运而生。
差分进化算法(differential evolution, DE)是一种基于群体差异的启发式随机搜索算法, 该算法是Storn 等[20]为求解切比雪夫多项式而提出的。因原理简单、受控参数少等特点, 在约束优化计算、聚类优化计算、非线性优化控制、神经网络优化、滤波器设计、阵列天线方向图综合及其他方面得到广泛应用[21]。基本差分进化算法采用“贪婪”选择操作, 经过变异与交叉操作后生成的实验个体xT与xit进行比较, 只有当xT的适应度较xit更优时才被选做子代, 否则, 直接将xit作为子代[22]。选择操作的方程为
$x_{i}^{t+1}=\left\{ \begin{align} & {{x}_{T}},\ \ f({{x}_{T}})<f(x_{i}^{t})\ , \\ & x_{i}^{t},\ \ f({{x}_{T}})\ge f(x_{i}^{t})\ , \\ \end{align} \right.$
式中, f(·)为个体的适应度值, 在求解约束优化问题时, 选择和比较个体时必须同时考虑目标函数和约束条件, 然而, 差分进化算法在进化的过程中不可避免地会产生进化停滞现象, 进而使种群容易发生早熟收敛。为了解决这一困难, 各种差分寻优策略应运而生, 但每种策略只是针对性不同, 如 DE/ rand/1 有利于保持种群的多样性, 而 DE/best/1 更强调寻优的方向性和收敛速度。两种差分策略也各有局限性。不同领域的学者开始研究引入各种算子策略对差分进化算法进行改进[23]。
为了克服差分进化算法中 DE/rand/1 和DE/ best/1 的局限性, 很多人根据研究内容的不同引入不同的算子。王立国等[24]利用遗传算法与蚁群算法相结合的手段来研究高光谱图像的波段选择问题, 既利用了遗传算法中以较快的方式寻求最优解的特点, 又利用了蚁群算法中编码简单、耗时少的优点, 提高了算法的搜索效率。本文则将改进差分进化算法与突变论扩展模型相结合, 在得到 10 个树种的材积-树高-胸径模型的同时, 缩短了计算时间, 也提高了模型的收敛速度。
试验场地选在内蒙古赤峰市喀喇沁旗旺业甸林场。喀喇沁旗土地总面积为 3050 km2, 其中耕地434666.7 hm2, 有林地 152000 hm2。属中温带大陆性季风气候, 春季风大干燥, 夏季多雨高温, 秋季霜冻较早, 冬季寒冷少雪, 四季分明。具体气候情况见表1。
表1 研究区域气候状况
Table1 Climatic conditions of the study area
按照径阶的不同, 随机取样测定旺业甸林场内1000 棵树, 包括红松、油松、落叶松、樟子松、五角枫、山杨、榆树、柞树、白桦和黑桦等 10 个树种, 每个树种随机抽取 100 棵, 不同树种按照树高、胸径生长的国际标准值取长势良好的树木。利用电子经纬仪和测树枪两种主要仪器, 对 1000 棵树的树高、地径、胸径进行测定, 并以此作为真值。同时, 随机抽取每个树种的边缘树种, 即非标准木的树种, 每个树种抽取100棵对所建立模型进行精度检验。
利用改进差分进化算法, 建立内蒙古赤峰市旺业甸林场1000棵树的10种不同径阶的树种材积模型, 如表 2 所示。由于突变论模型基础是 V(x)= x4+x2+x, 本研究的扩展模型是 V(x)=ax14+bx22+ cx3+d, 即在通过电子经纬仪等仪器测定树高 H、地径 D0 和材积 V 的情况下, 利用改进差分进化算法对扩展模型中的未知变量进行计算, 得到树高系数 a、胸径系数 b、地径系数 c 和常数项d的值, 运算至达到最佳收敛判断标准为止。从我国北方 10 个树种材积、树高、胸径及地径的相关性可以看出, 各树种模型的相关系数 R 均大于 0.9, 说明模型计算出的结果跟实测值相差不大, 即模型拟合精度较高, 同时说明我国北方 10 个常见树种的材积变化符合突变论的尖角模型理论, 即随着树高的增长及树木材积的变化, 在树形逐渐趋向三角形的顶点时, 树木的材积会有一个实质性的变化。
从图 4 可以看出, 随着树高的增长, 单木的材积逐渐增大, 尽管因树种的不同, 材积增量存在差异, 但整体上都呈上升的增长趋势。在树木的整个生长期内, 在不同的生长阶段长势并不相同, 但都是在生长初期(即幼苗期)苗木生长相对缓慢, 生长中期是苗木的速生期, 苗木生长迅速。不同树种树木的速生期开始时间不同, 曲线的第一个转折点即标记为苗木速生期的开始。图中实线代表单木的实际生长情况, 虚线代表模型模拟的情况。可见, 各树种的实际生长和模拟生长曲线的拟合度较好。同时, 表明利用突变论扩展模型来说明单木的生长变化是合理的。为了验证这一模型的精确度, 本试验随机抽取 10 个树种的非标准木对该模型进行外符合精度的检验。
图4 中国北方10个树种的模型预测值与实测值间的比较 Fig. 4 Comparison between predicted and measured values of 10 species in northern China
利用突变论的尖角模型理论建立的 10 个不同树种的模型, 在经过不同的迭代次数计算得出: 模型的内符合精度和相关系数都达到 90%以上(如表2 所示), 说明利用突变论的尖角模型理论研究不同阶段树木的单木材积是可行的, 也说明不同树种的单木材积在树高达到一定高度时符合突变论的尖角模型理论, 材积达到一定程度上的突变, 为了争夺更多的生长空间, 从而引发树种间的竞争。同时, 抽取 10 个树种的边缘树种(即各个树种的非标准木, 每个树种 100 棵), 对所建立模型进行外符合精度卡方检验, 结果如表3所示。
表2 基于改进差分进化算法的10种中国北方常见树种的材积突变模型及卡方检验
Table 2 Volume catastrophe model and chi-square test based on improved differential evolution algorithm of the 10 common tree species in northern China
表3 边缘树种对10个树种模型的精度检验
Table 3 External accord accuracy test of the 10 tree species model used edge species
利用改进差分进化算法对模型进行检验, 结果是不同的树种经过不同的迭代次数和迭代时间之后, 都在一定程度上达到最优化收敛。从表3可以看出, 10个树种模型检验的迭代次数都在250和340这个区间内, 计算到最优所需要的时间都很接近, 说明每个树种选取100棵边缘值进行检验, 各个模型的计算时间都相近。模型总体相对误差RS在[0.001, 0.05]区间内, 平均相对误差 E 在[-0.11, 0.02]区间内, 总体预估精度 P (即外符合精度)都大于 80%, 说明模型拟合较好, 也说明各树种的非标准木也适合本研究建立的材积-树高-胸径模型。通过上述研究表明, 基于突变论尖角模型所建立的材积-树高-胸径模型具有较高的计算精度, 为材积的计算和林分蓄积的计算提供了一定的理论和实践依据, 提高了林木材积数据获取的工作效率。
本文将突变论的尖角模型理论引入我国北方10 个常见树种材积模型中, 通过试验表明单木的生长也符合突变理论。即随着树木的生长, 在树高生长达到一定程度时, 材积将不再发生较大变化。在模型构建过程中, 为了解决树种多, 数据量庞大使得计算时间过长的问题, 本文结合改进差分进化算法使得模型在短时间内收敛到最优解, 并通过卡方检验, 将模型的内符合精度达到90%以上, 说明突变论的尖角模型理论适用于材积-树高-胸径模型, 即在林木达到一定的生长阶段, 随着树高的增长, 树木的胸径趋向于一个点, 材积在此时达到“突变”。另外, 通过测量每个边缘树种的树高、胸径、地径和材积来对各个模型进行外符合精度的检验, 检验结果为外符合精度全部大于80%, 达到模型的精度标准, 说明本试验所做的“突变”模型适用于我国北方常见10 个树种的标准木和非标准木的林木材积计算,为以后树种材积和树种间空间竞争的研究提供了一定的理论和实践依据。下一步将会继续结合树冠表面积等空间因子, 对树种间的竞争进行深入研究。
The authors have declared that no competing interests exist.
突变论在种间生态学中的应用 突变论模型在生态系统中的应用 关于常微分方程数学模型的建立分析 A stochastic catastrophe model using two-fluid model parameters to investi-gate traffic safety on urban arterials 奇点(含突变)理论的某些应用 Catastrophe theory and fisheries regulation Catastrophe theory applied to ecological systems Catastrophe theory in ecology: a critical review and an example of the butterfly catastrophe 基于突变论的储层边界尖点突变技术及应用研究 基于突变理论模型的卧龙湖湿地退化判别研究 基于蜱虫效应的Sgn函数突变模型 An index to assess South Africa vulnerability to sea-level rise 基于遥感及突变理论的生态环境脆弱性时空演变: 以黄河三角洲垦利县为例 基于神经网络的森林火灾危害程度预测研究 基于神经网络的 GPS 高程拟合方法优选及精度分析 不同氮磷比条件对外来种火炬树与本地种麻栎、荆条的种间关系的影响[D] 一种改进的遗传算法 基于生物入侵思想的自适应遗传算法优化 Differential evolution: a simple and efficient adaptive scheme for global optimization over continuous spaces 改进差分进化算法及其应用 一种改进的约束优化差分进化算法 遗传算法在最优化问题中的应用 结合遗传算法和蚁群算法的高光谱图像波段选择
/
, Haiying MAO
1 原理与方法
1.1 突变论模型
x为状态变量, u和v为控制变量, a点为突变点; 轨迹Ⅰ突然变化, 轨迹Ⅱ连续变化
1.2 遗传算法
1.3 差分进化算法
1.4 改进差分进化算法
2 试验与分析
2.1 试验地概况
地区 年均气温/℃ 年积温/℃ 极端低温/℃ 极端高温/℃ 无霜期/d 年日照/h 年均风速/(m·s-1) 年均降水/mm 赤峰市 0~7 2600~3800 -10 41 78~90 2700~3100 2.5 300~500 喀喇沁旗 3.5~7 2000~3200 -30.9 37.4 110~150 2913.3 2.8 591.2
2.2 试验材料
2.3 模型的建立
2.4 精度的分析
树种 模型 RMSE SSE R R2 DC C-S F 油松 Pinus tabuliformis. y=-40.671x14-2.975x22
+2.742x3+13.7530.0200 0.0064 0.9969 0.9938 0.9938 0.00881 2232.1520 樟子松 Pinus sylvestnis var. mongolica Litv. y=-5.048x14+9.821x22
-0.0002x3+0.00140.0044 0.0002 0.9995 0.9990 0.9990 0.0017 9134.0809 落叶松 Larix gmelinii (Ruprecht) Kuzeneva. y=-1.927x14+9.067x22
+2.754x3-0.0570.0520 0.0461 0.9919 0.9839 0.9839 0.1097 918.9520 红松 Pinus koraiensis Siebold et Zuccarini. y=-7.958x14+0.0001x22
+0.00032x3-0.04290.0090 0.0007 0.9936 0.9873 0.9873 0.0041 467.1430 五角枫 Acer elegantulum Fang et P. L. Chiu. y=-5.536x14+8.979x22
-1.837x3+4.4670.0015 1.3536 0.9998 0.0006 0.9996 0.0002 9952.2379 榆树 Ulmus pumila L. y=-4.199x14+0.0001x22
-0.0001x3+0.00030.0355 0.0151 0.9904 0.9809 0.9809 0.0289 514.8484 柞树 Xylosma racemosum. y=-6.343x14+7.258x22
+0.0005x3-0.04210.0152 0.0032 0.9945 0.9891 0.9891 0.0400 1085.7894 黑桦 Betula dahurica Pall. y=-3.544x14+9.346x22
+2.177x3-0.01830.0150 0.0033 0.9969 0.9939 0.9939 0.0135 1956.3780 白桦 Betula platyphylla Suk. y=-5.166x14-9.459x22
+0.0016x3-0.15940.0498 0.0447 0.9829 0.9662 0.9662 0.1472 457.7420 山杨 Populus davidiana. y=-2.507x14+7.583x22
+0.00032x3-0.07380.0700 0.0986 0.9913 0.9827 0.9827 -0.0509 1024.1950
树种 迭代次数 RS E P/% 油松-Pinus tabuliformis. 317 0.0200 -0.027 90.13 樟子松- Pinus sylvestnis var. mongolica Litv. 309 0.0436 0.008 97.21 落叶松-Larix gmelinii (Ruprecht) Kuzeneva. 338 0.0520 0.015 89.10 红松-Pinus koraiensis Siebold et Zuccarini. 296 0.0092 -0.113 87.12 五角枫-Acer elegantulum Fang et P. L. Chiu. 314 0.0015 0.008 88.17 榆树-Ulmus pumila L. 314 0.0355 0.017 89.21 柞树-Xylosma racemosum. 246 0.0151 -0.021 90.97 黑桦-Betula dahurica Pall. 280 0.0154 0.001 93.26 白桦-Betula platyphylla Suk. 267 0.0498 0.019 94.55 山杨-Populus davidiana. 294 0.0702 -0.007 96.78
3 结论与讨论
参考文献
[1]
[2]
[3]
[4]
[5]
[6]
[7]
[8]
[9]
[10]
[11]
[12]
[13]
[14]
[15]
[16]
[17]
[18]
[19]
[20]
[21]
[22]
[23]
[24]

〈
〉

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}