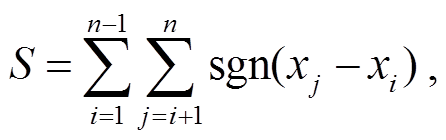 (1)
(1)
北京大学学报(自然科学版) 第60卷 第2期 2024年3月
Acta Scientiarum Naturalium Universitatis Pekinensis, Vol. 60, No. 2 (Mar. 2024)
doi: 10.13209/j.0479-8023.2024.008
国家自然科学基金长江水科学研究联合基金(U2240205)资助
收稿日期: 2023–03–12;
修回日期: 2023–03–23
摘要 根据洞庭湖城陵矶站 1990—2015 年月平均 COD, TN 和 TP 浓度的监测资料, 基于趋势分析方法, 系统地研究洞庭湖水体生源要素的年内变化和年际变化规律, 进一步揭示其化学计量比例特征, 在此基础上探讨生源要素浓度及结构对藻类生长的影响。结果表明, 多年平均 COD, TN 和 TP 浓度的年内变化规律相似, 均表现为非汛期>汛期; 多年变化趋势方面, 1990 年以来, 洞庭湖城陵矶站年平均 COD, TN 和 TP 浓度均呈现显著的上升趋势, 对应 C:N 和 C:P 随时间推移而下降, N:P 则表现为上升。洞庭湖城陵矶站水体 C:N:P 平均为 13:18:1, 冗余分析结果表明水体生源要素浓度及化学计量比会影响藻类生长, 其中 C:N, C:P 和 COD 起主导作用。研究时段内城陵矶站水体氮磷含量整体偏高, 已满足藻类生长需求, 为避免水华爆发等不良生态后果, 应控制外来氮磷输入, 保持湖体化学计量比的平衡。
关键词 洞庭湖; 生源要素; 化学计量比; 趋势分析
碳、氮和磷等生源要素是生物生长和生命活动必需的化学营养元素, 也是支持水体初级生产和维持水生系统结构与功能的保障。淡水系统中碳、氮和磷元素的生物地球化学循环通过初级生产过程相互耦合, 氮磷并不直接参与固碳过程, 但初级生产者需要它们来合成核苷酸、蛋白质和磷脂等生物大分子。湖泊生源要素对维持湖泊生态系统的稳定和健康至关重要, 不仅影响藻类群落的相对丰度与结构变化, 还会通过能量流动和物质循环改变食物网的结构[1–3]。同时, 湖泊是全球营养物质的重要储库, 可以有效地滞留和去除水体中的营养元素, 减轻人为输入的营养物质继续向下游生态系统输出的风险[4–5]。湖泊生源要素是湖泊生态系统健康状况的重要参数, 对湖泊的水资源管理和生态保护具有重要的指示作用。
20 世纪 80 年代末以来, 随着人口扩张和工业化, 全球人类活动导致生物圈的氮磷剧增, 且 N:P从 19 增加到 32, 明显高于其他生态系统的平均值(海洋为 15~16, 土壤为 16~22, 生物体为 4~30)[6–7]。过量的氮磷输入使得营养物质的浓度增加, 引起湖泊富营养化, 造成大型水生植物多样性锐减、生态系统结构和功能改变等一系列的后果[8–10]。然而, 富营养化也具有积极的作用, 湖泊的有机碳埋藏速率随着生产力的提高而增加, 湖泊作为潜在碳汇的重要性值得进一步研究评估[11]。湖泊内正常的营养物质循环是维持生态健康的必要条件, 主要包括源过程(如外源负荷、沉积物释放、大气沉降和生物固氮等)、去除过程(如湖体出流、沉积埋藏和反硝化等)以及湖体内的迁移转化过程(如微生物分解和硝化作用等)[5]。大量氮磷的进入会导致湖体内营养物质循环过程发生变化, 湖泊水体中碳、氮和磷元素的相对比例和结构也因此而偏离原有的平衡[12]。研究表明, 全球湖泊的氮磷比在时间尺度上存在明显的上升趋势[4]。但是, 目前对湖泊内生源要素碳、氮和磷的浓度和组成变化规律及其造成的富营养化和生物多样性丧失等生态效应的了解十分 有限。
洞庭湖是中国第二大淡水湖泊, 位于湖南省中南部, 具有丰富的生物多样性, 支持着附近区域经济社会的发展[12–14]。然而, 在流域社会经济发展与资源开发利用的胁迫与压力之下, 洞庭湖水质恶化、富营养化风险加剧等问题逐步凸显, 水体氮磷含量超标, 局部水域(大小西湖)富营养化现象突 出[15], 给洞庭湖水生态环境健康及流域持续发展造成严重威胁。目前关于洞庭湖氮磷含量分布和季节变化特征已有相关研究, 对生态效应主要关注富营养化和水华风险[15–16], 对碳元素及碳氮磷化学计量比的时空分布特征了解甚少。因此, 本文对洞庭湖城陵矶站生源要素浓度的年内变化、年际变化趋势及化学计量特征进行深入的分析, 揭示藻类生长对生源要素的响应, 以期为洞庭湖生态系统保护和流域生态经济可持续发展提供科学支撑。
洞庭湖是我国五大淡水湖之一, 也是长江中下游重要的调蓄湖泊之一[12], 位于 E110°40′—113°10′, N28°30′—30°20′。湖体呈近似“U”字型, 流域面积为 26.28 万 km2, 占长江流域总面积的14.6%, 多年年均降水量约为 1400mm, 多年平均年径流量为 3.1×1011m3, 地表水资源在长江流域各水系中居首位[17]。洞庭湖水系复杂, 北连长江, 南纳洞庭“四水”(湘江、资水、沅江和澧水), 入湖水体主要由“四水”和长江“三口”(松滋口、太平口和藕池口)三部分组成, 经洞庭湖调蓄后在城陵矶(七里山)注入长江, 是一个典型的吞吐调蓄型湖泊[13–14]。城陵矶(七里山)水文站位于 E113°08′, N29°25′, 是洞庭湖唯一出口河段水文控制站, 被誉为洞庭湖及长江流域水情“晴雨表”, 同时也是洞庭湖水质监测的国控断面[18]。
城陵矶(七里山)水文站是洞庭湖的主要出口, 其水质情况代表洞庭湖出口水质, 反映湖体内营养物质循环过程和外来污染输入对洞庭湖水质的综合作用。同时, 洞庭湖是通江湖泊, 城陵矶(七里山)水文站是“江湖”的交汇点, 下游约 4km 为长江荆江与洞庭湖出流汇合口, 其水质直接对长江水资源的战略安全产生影响。因此, 本文选取洞庭湖城陵矶(七里山)水文站 1990—2015 年共 25 年的逐月平均COD, TN, NO3−, NH4+及 TP 水质监测数据(来自长江水利委员会), 分析生源要素分布特征。此外, 使用城陵矶站的藻类细胞密度数据(来自国家地球系统科学数据中心 http://www.geodata.cn/, 采样时间为2012—2014 年 3, 6, 9 和 12 月, 共 12 组), 通过冗余分析, 揭示洞庭湖浮游植物对水体生源物质浓度及其化学计量比的响应。
通过 Mann-Kendall 趋势检验, 明确洞庭湖碳氮磷生源要素浓度及化学计量比的变化趋势。分别对春季(3—5 月)、夏季(6—8 月)、秋季(9—11 月)、冬季(12—次年 2 月)、年平均生源要素浓度及碳氮磷化学计量比序列进行趋势检验, 以 Sen 斜率作为趋势值, 代表趋势变化的大小。
对样本数为 n 的时间序列 x1, x2, …, xn, M-K检验统计量 S 的计算方式[19]如下:
(1)
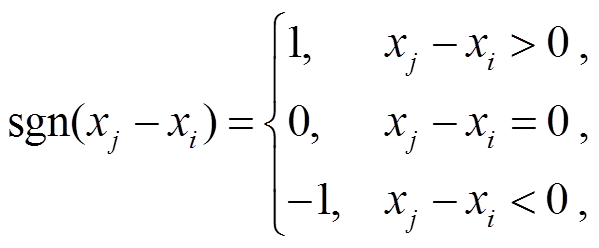 (2)
(2)
其中, n 为样本数量, xj和 xi分别为时间序列第 j 个和第 i 个样本(j>i), S 代表时间序列内各样本值大于之前时刻样本值的个数。N>10 时, 秩序列 S 的方差Var(S)为
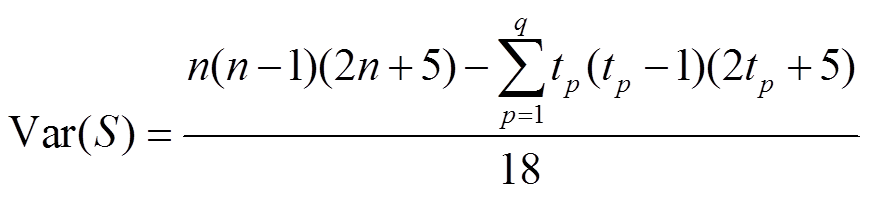 ,(3)
,(3)
其中, q 为时间序列中样本量数值相同的个数, tp 为第 p 组数值相同的样本量的个数。标准正态检验统计量 Z 的计算公式如下:
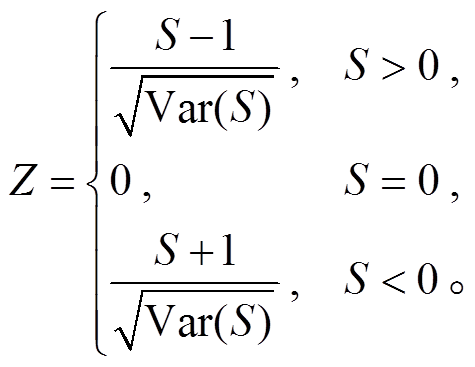 (4)
(4)
Z>0 表明序列呈上升趋势, Z<0 表明序列呈下降趋势。若|Z|>Z1−α/2, 表明时间序列存在显著的变化趋势, 其中 Z1−α/2 是标准正态偏差, α 为显著性水平。
Sen 斜率的计算方法[20]为
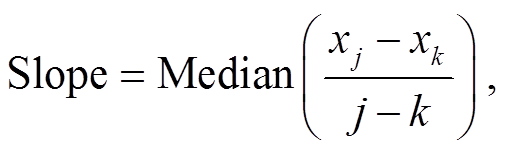 (5)
(5)
式(5)中, xj 和 xk 分别是 j 和 k 时刻的时间序列值, Slope 表示 Sen 斜率, Median 为中位数函数。Sen 斜率为正值表示时间序列呈现上升趋势, Sen 斜率为负值表示时间序列呈现下降趋势。
使用 R 语言进行冗余分析(redundancy analysis, RDA), 揭示水体中 COD, TN, TP 浓度及化学计量比与藻类生物量之间的关联。在此之前, 需要进行消除趋势对应分析(detrended correspondence analysis, DCA), 确定排序轴的梯度长度, 据此选择排序分析模型。通常, 排序轴梯度长度最大值大于 4 时, 选择典范对应分析(canonical correspondence analysis, CCA); 排序轴梯度长度最大值在 3~4 之间时, 选择RDA 和 CCA 均可; 排序轴梯度长度最大值小于 3时, 选择选择 RDA。经检验, 4 个排序轴中梯度长度最大值为 2.10, 因此采用RDA分析来反映洞庭湖浮游植物对生源要素的响应是可靠的。
图 1 显示洞庭湖城陵矶水文站 1990—2015 年多年平均 COD, TN 和 TP 浓度的年内变化特征。城陵矶水文站 1—12 月平均 COD 在 2.28~3.10mg/L 之间波动, 1—9 月逐渐下降, 10—12 月逐渐回升, 最低月平均 COD 出现在 9 月(2.28±0.48mg/L), 最高月平均COD 出现在 1 月(3.10±0.66mg/L)。对于 TN, 城陵矶水文站的多年月平均值变化范围为 1.32~2.02mg/L, 其年内变化规律与 COD 相同, 最低 TN 浓度值出现在 8 月。城陵矶水文站多年月平均 TP 浓度的峰值为 0.12mg/L, 谷值为 0.07mg/L, 与 COD 和TN 的年内变化规律不同, TP 浓度没有逐渐递增或递减的趋势, 而是呈现汛期(5 月—10 月)浓度整体低于非汛期(11 月—次年 4 月)的规律, 汛期或非汛期内各月份之间则无明显差异。
洞庭湖生源要素的季节分布规律与长江下游的鄱阳湖类似, 鄱阳湖 TN 和 TP 含量也呈非汛期>汛期的趋势[21]。非汛期湖泊径流量少, 水环境容量有限, 对氮磷和有机污染的净化能力弱。汛期雨水充沛, 洞庭湖上游来水较多, 大量农田化肥及生活污水随雨水汇入河槽, 形成径流汇入湖体, 但由于流量的稀释效应, 汛期 COD, TN 和 TP 的浓度反而降低[22]。
图 2 显示 1990—2015 年城陵矶水文站COD, TN和 TP 浓度的年际变化趋势。COD 多年年平均值为2.65mg/L, 1990—2015 年上升趋势显著(P<0.05), 从1.68mg/L 增加至 3.24mg/L, 但 2010 年后有明显的下降趋势。TN 浓度在 1998 年之前出现大幅波动, 此后呈上升趋势(P<0.001), 2010 年达到峰值 2.44mg/L, 2011—2015 年略有下降。TP 浓度的波动范围在 0.02~0.14mg/L 之间, 平均 0.09mg/L, 在 2003 年之前上升趋势显著(P<0.01), 2003 年后存在下降趋势。
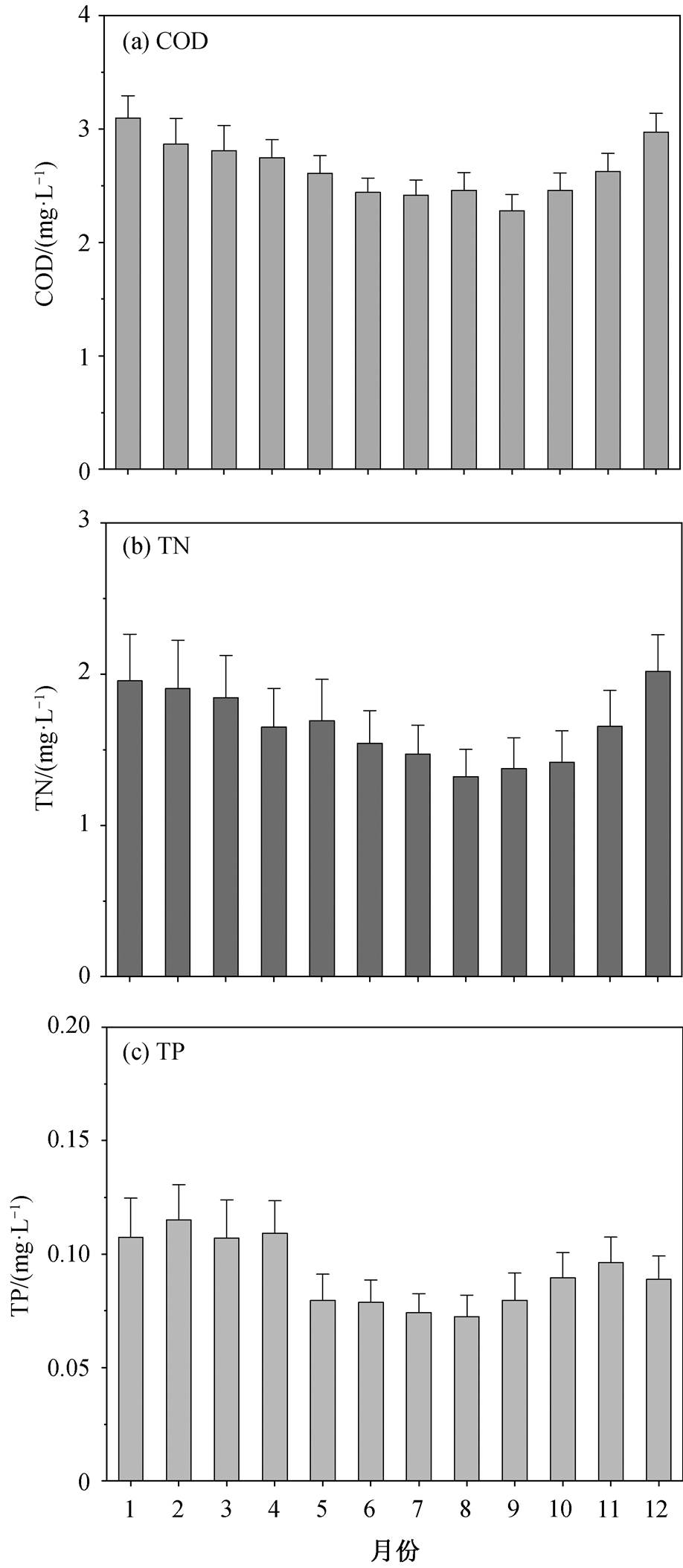
图1 城陵矶水文站1990—2015年多年月平均COD, TN和TP浓度的年内变化
Fig. 1 Intra-annual variation of average COD, TN and TP concentrations at Chenglingji hydrological station from 1990 to 2015
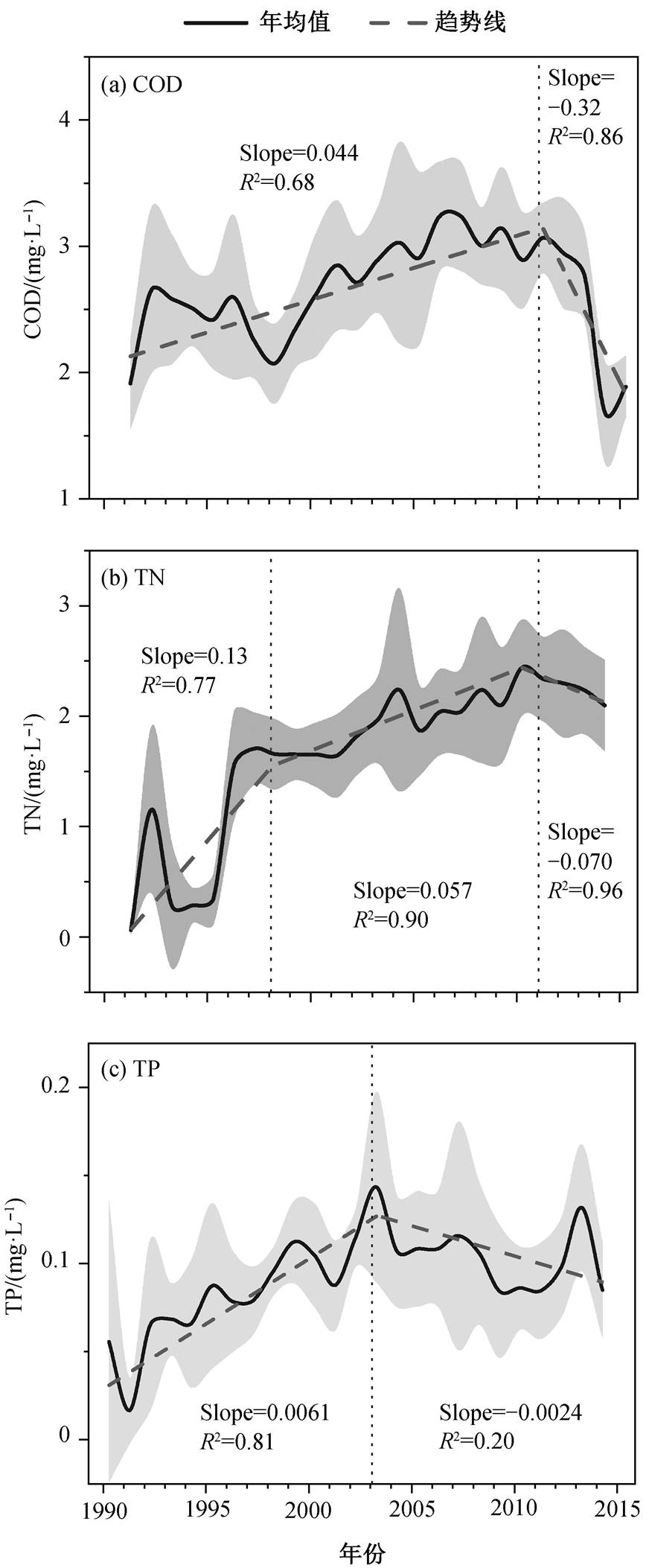
图2 城陵矶水文站 1990—2015 年 COD, TN 和 TP 浓度的年际变化趋势
Fig. 2 Annual trends of COD, TN and TP concentrations at Chenglingji hydrological station from 1990 to 2015
研究时段内, 各季节生源要素浓度的变化趋势与全年均值保持一致(表 1)。对于 COD, 夏季(P<0.05)和秋季(P<0.01)增势显著, 趋势变化值也最大, 分别为 0.040 和 0.030。春秋两季 TP 浓度显著上升, 尤其是春季, Sen 斜率达到 0.0034。TN 浓度在各季节均保持显著上升, 其中春季的趋势变化值最高(0.090)。
表1 1990—2015年城陵矶水文站各季节COD, TN和TP浓度的趋势分析
Table 1 Trend analysis of COD, TN and TP concentrations in different seasons at Chenglingji hydrological station from 1990 to 2015
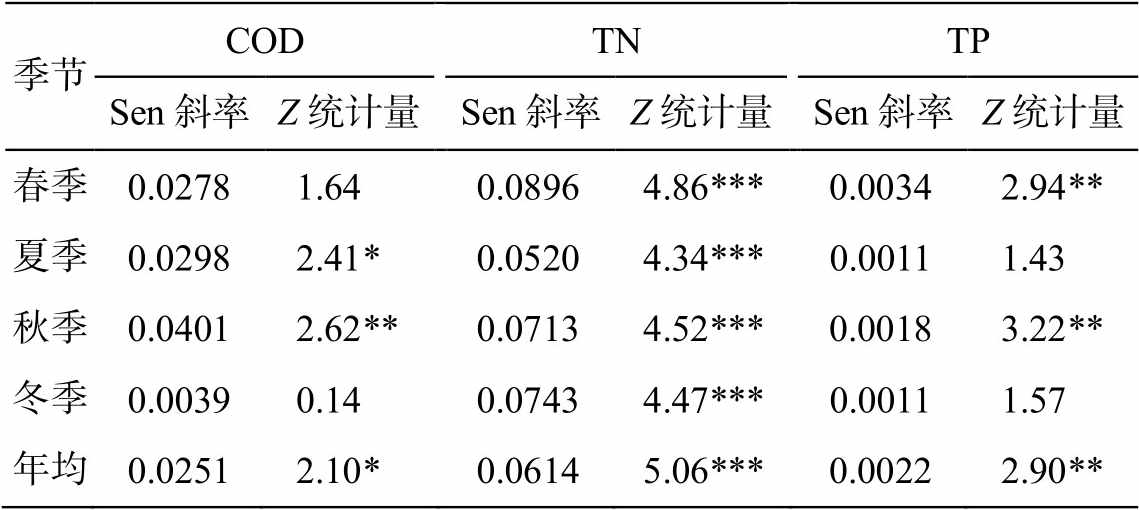
季节CODTNTP Sen斜率Z统计量Sen斜率Z统计量Sen斜率Z统计量 春季0.02781.640.08964.86***0.00342.94** 夏季0.02982.41*0.05204.34***0.00111.43 秋季0.04012.62**0.07134.52***0.00183.22** 冬季0.00390.140.07434.47***0.00111.57 年均0.02512.10*0.06145.06***0.00222.90**
注: *P<0.05, **P<0.01, ***P<0.001。
上述结果表明, 洞庭湖水体生源要素浓度在过去几十年间显著增加, 水质逐渐恶化, Geng 等[23]关于洞庭湖水质评价的研究也得出同样的结论。污水排放是洞庭湖污染加剧的主要原因之一, 1996 年至2015 年, 洞庭湖地区的污水排放增长率高达 32%, 很多造纸、纺织和化工等重污染工业的污水未经处理, 直排入湖, 2006 年的排放量达到 7181 万吨[24–25]。污染物的大量流入导致洞庭湖生源要素浓度增加, 水质随之恶化。其次, 洞庭湖地区是中国重要的农业生产基地, 大量的化肥用于粮食增产, 未被植物吸收的过剩氮磷流失到湖体中, 从而增加 TN 和 TP浓度[24]。第三, 1996 年至 2010 年的平均污水处理率仅为 36%, 远低于全国平均水平, 未能有效地减少洞庭湖的外来营养物质输入[25]。
值得注意的是, COD 和 TN 浓度 2010 年前后达到峰值后表现出下降趋势, 这与当地政府出台实施的环保政策密不可分。2009 年发布《湖南省洞庭湖水环境综合治理规划实施方案》后, 城市污水和工业废水处理设施得到完善, 农村生活污水处理也得到推广。洞庭湖地区污水处理率在 2010 年迅速增加到 74%, 而 1999 年至 2008 年期间仅为 17%~ 58%[24]。这些措施有效地限制了洞庭湖的外源营养物质输入, COD 和 TN 浓度均有所下降, 缓解了水体营养负荷高和水质恶化问题。然而, 2010 年之后, TN 浓度仍然保持较高水平, 这与农业生产过程中持续施用氮肥密切相关, 1996 年至 2015 年, 洞庭湖地区化肥施用量增长 59%。此外, 据报道, 2010 年洞庭湖地区的氮肥施用率高达 290.8kg/hm2, 而氮肥利用率约为 37%, 大量的氮肥流失导致洞庭湖水体 TN 浓度高居不下[26], 水生态健康状况令人堪忧。除污水排放外, 水文条件的变化也会显著地影响洞庭湖生源物质的动态平衡, 增加水环境风险。三峡大坝的运行对洞庭湖的水文条件产生重大影响, 洞庭湖的平均径流量和泥沙通量分别从 2.48× 1011m3/a和 88.28×106t/a 降至 2.09×1011m3/a和 16.80 ×106t/a[24]。长江水环境中总磷以颗粒磷为主, 泥沙是颗粒磷的主要载体。长江贡献了洞庭湖80%的入湖沙量, 也是洞庭湖总磷的主要来源, 然而上游三峡大坝导致入湖沙量减少 81.0%, 从而造成洞庭湖总磷浓度下降。之前的研究也证实, 三峡水库拦截了入库水体中 42.5%的磷, 其中颗粒磷滞留量占81.5%。因此, 大量磷滞留在长江上游, 输送至中下游的磷通量大幅度减少[27]。
洞庭湖水体碳氮磷化学计量比的分布情况见图 3。C:N 比值介于 0.3~27.6 之间, 平均 0.7; C:P 波动幅度很大, 最低值为 4.8, 最高值为 647.5, 均值为13.1。N:P 在 0.5~792.2 之间变化, 平均为 17.8。总体而言, 洞庭湖水体 C:N:P 平均值为 13:18:1, N:P 与经典的 Redfield 值(16:1)相近, 而 C:P 与 Redfield 值(106:1)差距较大[28]。据报道, 长江中下游湖泊中不同营养类型的湖泊 C:N:P 有所不同, 富营养湖泊C:N:P 平均值为 88:58:1, 中富营养湖泊为 73:35:1, 中营养湖泊为 44:22:1[26]。洞庭湖全湖处于中营养水平, 局部处于富营养水平, 本研究中洞庭湖 N:P与中营养湖泊相近, 但 C:P 和 C:N 显著低于其他长江中下游湖泊, 可能与洞庭湖氮磷污染严重有关, 研究时段内湖体氮和磷浓度显著上升, 并且增长率远高于 COD。已有研究证实, 湖泊外部营养物质输入和内部营养循环过程的共同作用影响着湖体化学计量平衡。全球 90%以上湖泊的流出水体 N:P 值远高于流入水体[4], 说明湖泊内部营养循环对水体化学计量平衡进行了重塑, 包括氮磷沉积释放、固氮作用和反硝化作用等。但是, 总体而言, 外部营养物质输入对湖体化学计量平衡具有更大的影响[4]。
图 4 显示1995—2015年城陵矶水文站碳氮磷化学计量比的年际变化趋势及趋势检验结果。与 20 世纪末相比, 洞庭湖水体的 C:N:P 已经发生巨大的变化, 逐渐偏离原有平衡。1995—2015 年 C:N 下降趋势显著(P<0.01), 从 0.78 减至 0.35, 期间趋势变化大小为−0.024; 研究时段内 C:P 处于波动状态, 趋势变化不明显(P>0.05), 多年平均值为 13.3, 但2010 年后出现下降趋势。与 C:N 和 C:P 相反, 洞庭湖水体 N:P 呈现显著上升趋势(P<0.01), 从 1995 年的 5.5 增至 2015 年的 27.6, 净增长 4.0 倍, 增加趋势值为 0.802。M-K 趋势检验结果(图 5)显示, 各季节水体 C:N:P 变化趋势与全年均值保持一致, C:N 随时间推移而显著下降, N:P 表现出显著上升趋势。单一的生源要素浓度(COD, TN 和 TP)不平衡的增长是湖体化学计量比失衡的主要原因。1990—2015 年, TN 和 TP 浓度的增长率分别为 112%和 81.5%, 而COD 增长率仅为 18.3%, 约为 TN 和 TP 浓度增长率的 1/6 和 1/4~1/5。由此可见, TN浓度相对更高的增长率是湖体 C:N 持续下降, N:P 持续上升的驱动 因素。
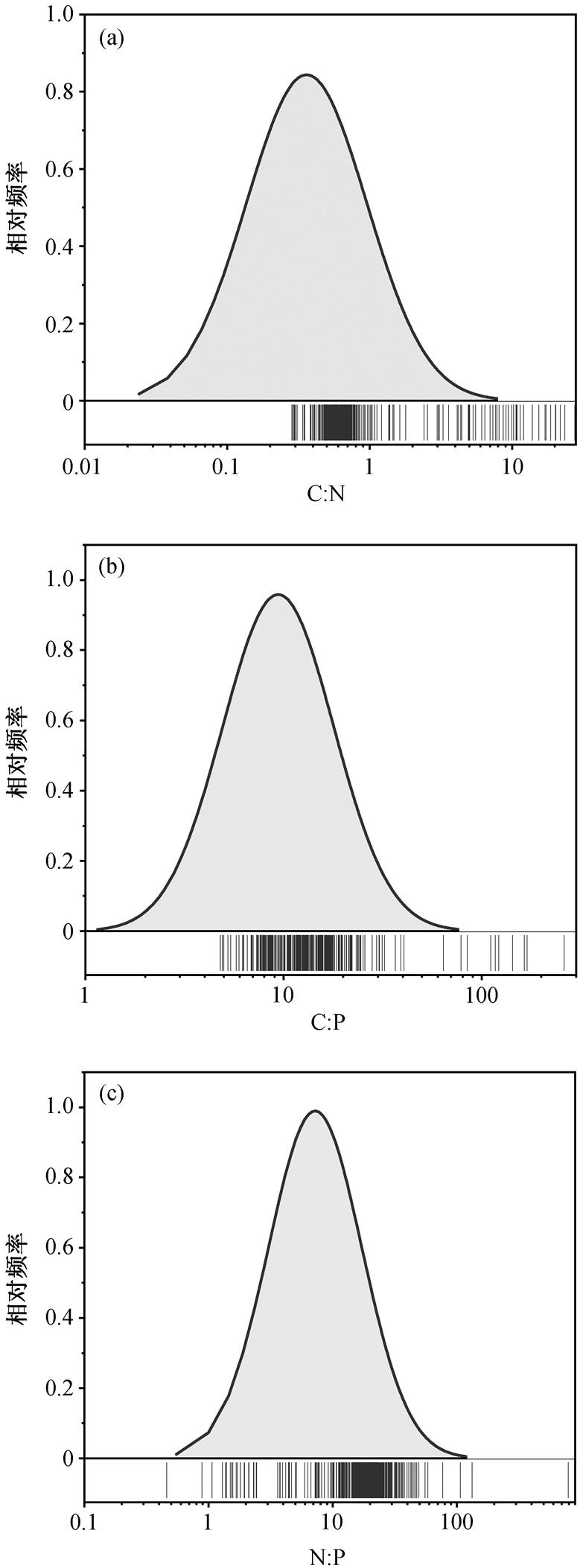
图3 洞庭湖水体碳氮磷化学计量比分布情况
Fig. 3 Stoichiometry of carbon, nitrogen, and phosphorus in Dongting Lake
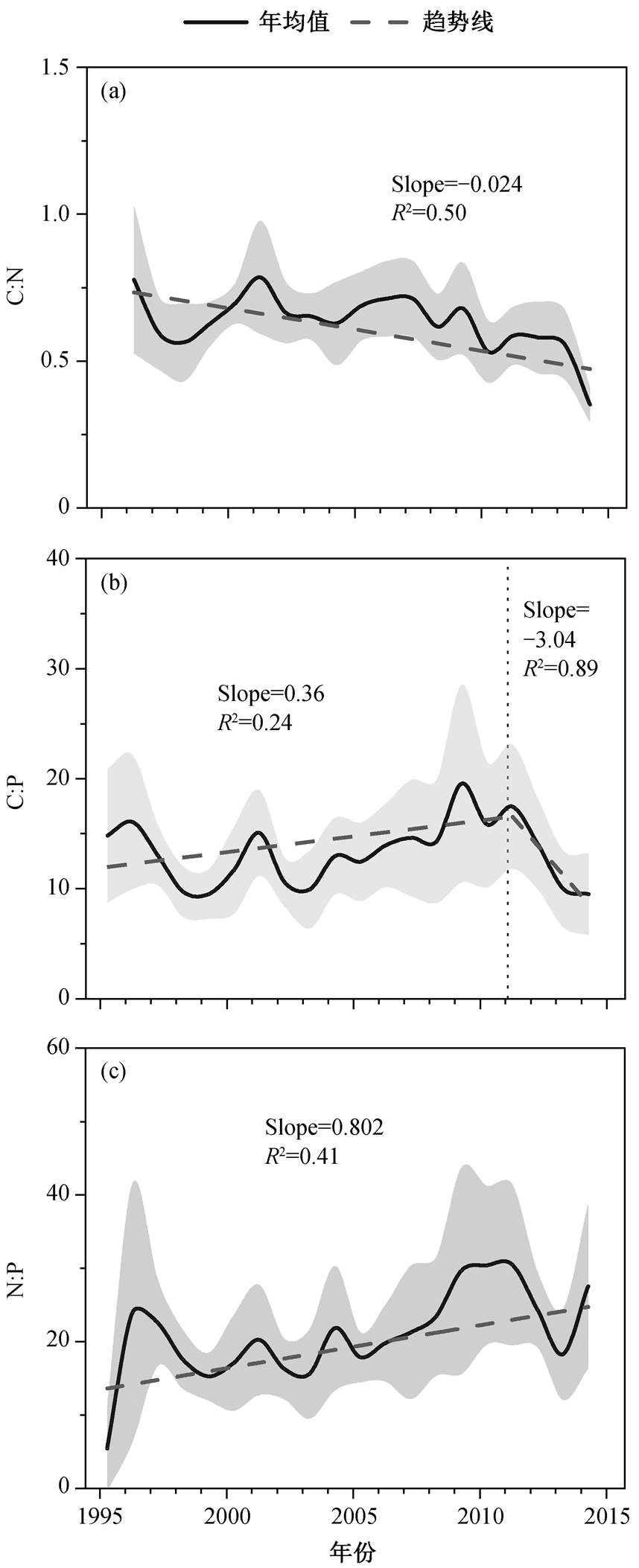
图4 城陵矶水文站 1995—2015 年 C:N, C:P 和 N:P的年际变化趋势
Fig. 4 Annual trends of C:N, C:P and N:P at Chenglingji hydrological station from 1995 to 2015
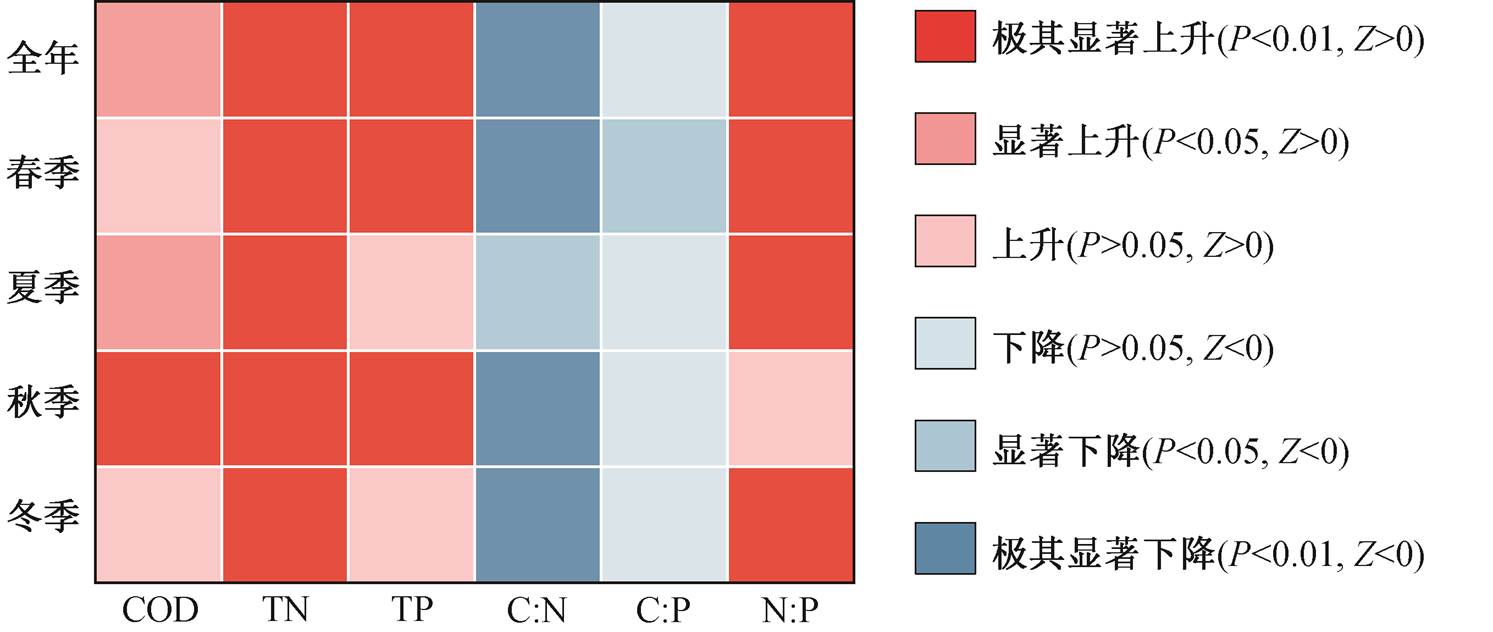
图5 洞庭湖生源要素浓度和化学计量比的Mann-Kendall趋势检验结果
Fig. 5 Mann-Kendall test results for concentrations and stoichiometry of carbon, nitrogen and phosphorus in Dongting Lake
生源要素比例结构的变化趋势反映洞庭湖水体化学计量比正逐渐失衡, 可能造成水质恶化、生物多样性锐减和生态系统生产力下降等不良的生态后果, 值得引起重视。为揭示生源要素浓度及组成对藻类生长的影响, 本文进一步通过冗余分析来解释城陵矶站碳氮磷浓度及化学计量比对藻类生物量的影响程度。冗余分析结果(图 6)显示, COD, TN, TP, NH4+, NO3−, C:N, C:P 和 N:P 显著影响藻细胞密度, 对 RDA 前两轴的解释度之和为 39.01%, R2 依次是0.396, 0.016, 0.101, 0.264, 0.186, 0.462, 0.421 和0.114。可见, 在各生源要素浓度及其化学计量比中, C:N, C:P 和 COD 是影响洞庭湖城陵矶站藻类密度的主要因子。
生源要素是藻类生长必需的养分, 藻类细胞分裂过程中会按照一定的比例吸收碳、氮和磷等营养元素。生源要素的比例和结构对藻类生长有显著的影响[29]。一些学者提出水体中营养盐限制性划分标准[30–31]: 1) N:P<20, 水体中浮游植物生长受氮限制, 有可能发生生物固氮作用来调节 N:P; 2) N:P> 50, 磷将成为浮游植物生长的限制性营养因子, 较低的磷含量水平可能使浮游植物对氮的有机合成受到抑制; 3) N:P 介于 20~50 之间, 适宜浮游植物的生长, 会加速浮游植物对氮元素有机合成的生态过程, 促进浮游植物爆发性生长。洞庭湖水体 N:P 的平均值为 17.8, 表明相较于磷, 氮含量水平较低, 氮元素是洞庭湖水体藻类生长的限制性营养因子。但是, 在营养水平较高(TN>0.2mg/L, TP>0.02mg/L)的情况下, 水体氮磷营养盐对藻类的限制作用会减弱。当前洞庭湖水体的 TN 和 TP 含量已满足藻类生长需求, 因此有机碳和其他因素(如水下光照条件、水温和水动力条件)对藻类生长繁殖和水华暴发的影响更大[18,32]。
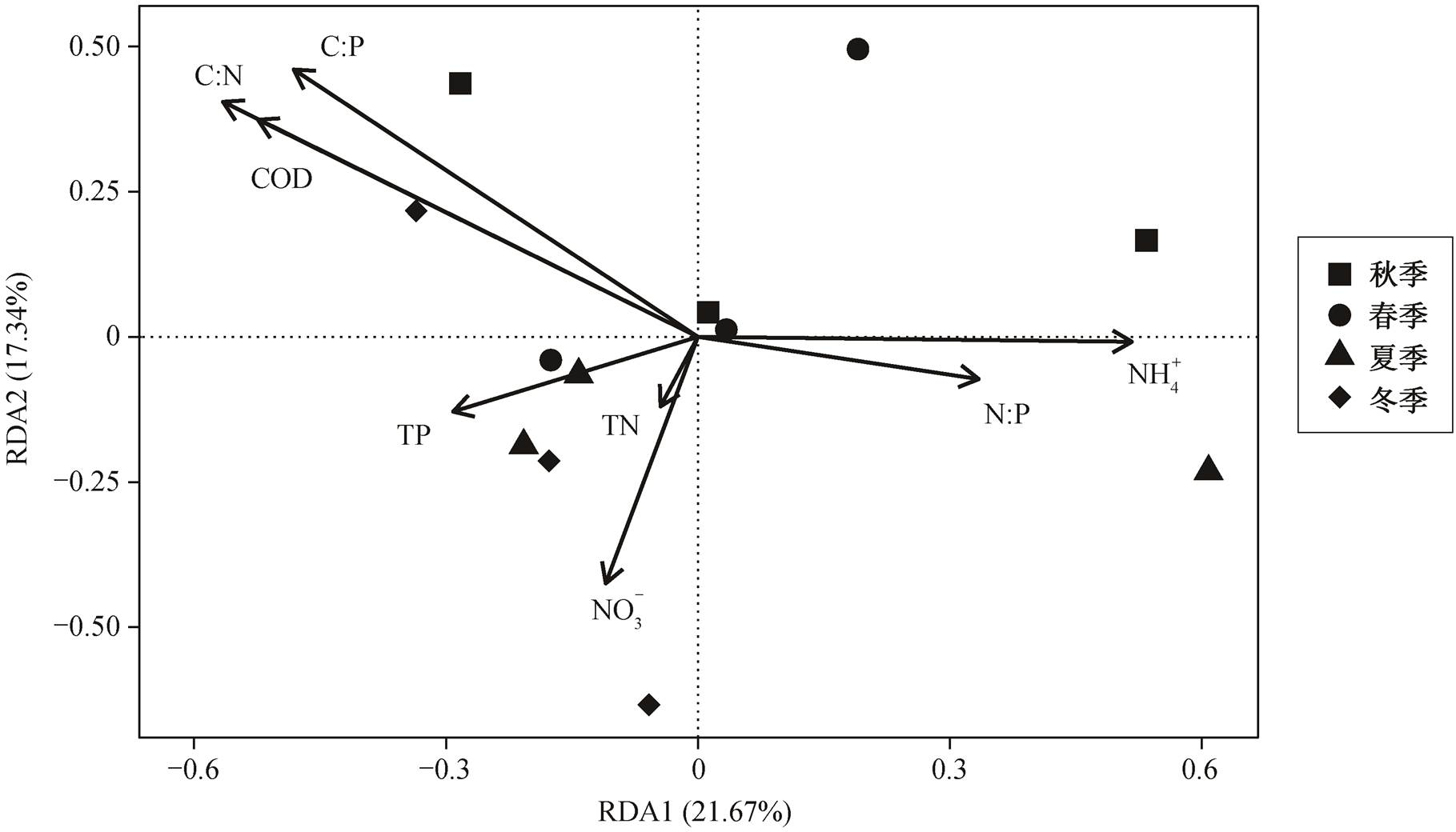
图6 冗余分析显示藻类生物量与生源要素浓度及化学计量比的关系
Fig. 6 Redundancy analysis showing algae biomass in relation to nutrient concentrations and stoichiometry
2008 年后, 洞庭湖浮游植物数量急剧上升, 尤其以东洞庭湖数量为最高, 达到 13.7×105 L−1, 远高于 20世纪 90 年代初的水平(0.9×104~2.45×104 L−1)。2008 年东洞庭湖的大小西湖及附近连通水域首次出现水华, 随后水华爆发面积逐年增加, 最大面积接近东洞庭湖 1/3 水域的面积[32]。2008 年以前, 洞庭湖并未出现大面积水华爆发现象, 全湖一直处于中营养水平, 局部地区为富营养水平, 原因在于洞庭湖属于过水型湖泊, 径流量大, 泥沙含量高, 水循环周期短, 这一独特的水文情势对富营养化发展和水华爆发风险有一定的抑制作用[33]。但是, 三峡水库运行后, 湖体流速减缓, 水力停留时间增加, 泥沙含量降低, 透明度增大, 同时加剧了营养物质富集, 局部水域水动力条件和水下光照条件变得更有利于藻类生长, 尤其是流速较低的东洞庭湖[34]。因此, 降低水体氮磷含量, 保持水体营养结构平衡是避免富营养化和水华的关键措施, 应当有效地控制流域氮磷负荷输入, 同时也要关注水文情势变化对水环境的影响[34]。
本文通过对洞庭湖城陵矶站 1990—2015 年生源要素浓度和碳氮磷化学计量比序列数据的趋势分析, 以及对营养盐与藻细胞密度的关联分析, 得到如下结论。
1)洞庭湖城陵矶站多年平均 COD, TN 和 TP 含量有相似的年内变化规律, 表现为非汛期>汛期。COD 和 TN 含量在 1—8 月逐渐下降, 9—12 月逐渐回升, TP 含量则无递增或递减现象。
2) 1990—2015 年洞庭湖城陵矶站 COD, TN 和TP 含量呈现上升趋势, 趋势变化率分别为 0.025, 0.061 和 0.002。各季节的生源要素变化趋势与年均值保持一致。
3)洞庭湖城陵矶站水体的 C:N:P 平均值为 13: 18:1, 1995—2015 年水体的 C:N 和 C:P 值随时间推移而下降, 而 N:P 值表现出上升趋势, 主要原因在于生源要素浓度不平衡的增长, TN 浓度增长率远高于 COD 和 TP。
4)洞庭湖水体生源要素浓度及化学计量比会影响藻类生长, 其中 C:N, C:P 和 COD 对城陵矶站藻类生长的影响起主导作用。研究时段内, 水体氮磷含量较高, 已满足藻类生长需求, 为避免水华风险, 应注意管理流域营养物质输入, 尤其是氮磷, 同时保持营养盐结构的平衡。
参考文献
[1] 同萌. 长江流域水库及河口沉积物生源要素对比研究[D]. 上海: 华东师范大学, 2020
[2] Feng W, Yang F, Zhang C, et al. Composition charac-terization and biotransformation of dissolved, particu-late and algae organic phosphorus in eutrophic lakes. Environmental Pollution, 2020, 265: 114838
[3] 严维霞, 吕路. 水库运行对生源要素生物地化过程的影响及生态效应. 环境科技, 2017, 30(4): 68–71
[4] Wu Z, Li J, Sun Y, et al. Imbalance of global nutrient cycles exacerbated by the greater retention of phos-phorus over nitrogen in lakes. Nature Geoscience, 2022, 15: 464–468
[5] 吴桢, 吴思枫, 刘永, 等. 湖泊氮磷循环的关键过程与定量识别方法. 北京大学学报(自然科学版), 2018, 54(1): 218–228
[6] Penuelas J, Janssens I A, Ciais P, et al. Anthropogenic global shifts in biospheric N and P concentrations and ratios and their impacts on biodiversity, ecosystem productivity, food security, and human health. Global Change Biology, 2020, 26(4): 1962–1985
[7] Peñuelas J, Sardans J. The global nitrogen-phosphorus imbalance. Science, 2022, 375: 266–267
[8] Schindler D W, Carpenter S R, Chapra S C, et al. Reducing phosphorus to curb lake eutrophication is a success. Environmental Science and Technology, 2016, 50(17): 8923–8929
[9] Glibert P M. Eutrophication, harmful algae and bio-diversity — challenging paradigms in a world of complex nutrient changes. Marine Pollution Bulletin, 2017, 124(2): 591–606
[10] Glibert P M, Maranger R, Sobota D J, et al. The Haber Bosch-harmful algal bloom (HB-HAB) link. Environ-mental Research Letters, 2014, 9(10): 105001
[11] Liu Q, Liang Y, Cai W J, et al. Changing riverine organic C:N ratios along the Pearl River: Implications for estuarine and coastal carbon cycles. Science of The Total Environment, 2020, 709: 136052
[12] 白倩倩, 梁恩航, 王婷, 等. 洞庭湖表层水温变化特征及其对气候变化的响应. 北京大学学报(自然科学版), 2022, 58(2): 345–353
[13] 赵子豪. 洞庭湖水文条件变化对水华发生风险影响模拟研究[D]. 南昌: 南昌大学, 2022
[14] 龚川. 城陵矶水位波动下东洞庭湖大小西湖水华形成过程研究[D]. 武汉: 湖北工业大学, 2020
[15] 曹萌萌, 毛克彪, 严毅博, 等. 基于 MODIS 数据的洞庭湖水体和水华时空变化研究. 中国环境科学, 2019, 39(6): 2523–2531
[16] Wang L, Liang T. Distribution patterns and dynamics of phosphorus forms in the overlying water and sedi-ment of Dongting Lake. Journal of Great Lakes Re-search, 2016, 42(3): 565–570
[17] 符哲, 郭晶, 黄代中, 等. 洞庭湖的富营养演变特征及影响因素. 环境化学, 2022, 41(8): 2636–2645
[18] 王婷, 王坤, 王丽婧, 等. 三峡工程运行对洞庭湖水环境及富营养化风险影响评述. 环境科学研究, 2018, 31(1): 15–24
[19] Yue S, Wang C Y. The Mann-Kendall test modified by effective sample size to detect trend in serially corre-lated hydrological series. Water Resources Manage-ment, 2004, 18: 201–218
[20] Güçlü Y S. Improved visualization for trend analysis by comparing with classical Mann-Kendall test and ITA. Journal of Hydrology, 2020, 584: 124674
[21] 陈波. 鄱阳湖水体氮磷污染研究进展. 江西师范大学学报(自然科学版), 2016, 40(4): 437–441
[22] 黄代中, 李芬芳, 欧阳美凤, 等. 洞庭湖不同形态氮、磷和叶绿素a浓度的时空分布特征. 生态环境学报, 2019, 28(8): 1674–1682
[23] Geng M, Wang K, Yang N, et al. Evaluation and variation trends analysis of water quality in response to water regime changes in a typical river-connected lake (Dongting Lake), China. Environmental Pollution, 2021, 268: 115761
[24] Yin G, Liu L, Jiang X. The sustainable arable land use pattern under the tradeoff of agricultural production, economic development, and ecological protection — an analysis of Dongting Lake basin, China. Environ-mental Science and Pollution Research, 2017, 24(32): 25329–25345
[25] Ji X H, Zheng S X, Shi L H, et al. Systematic studies of nitrogen loss from paddy soils through leaching in the dongting lake area of China. Pedosphere, 2011, 21(6): 753–762
[26] 秦伯强, 高光, 朱广伟, 等. 湖泊富营养化及其生态系统响应. 科学通报, 2013, 58(10): 855–864
[27] Bao L, Li X, Cheng P. Phosphorus retention along a typical urban landscape river with a series of rubber dams. Journal of Environmental Management, 2018, 228: 55–64
[28] Redfield A C. The biological control of chemical factors in the environment. Science Progress, 1960, 11: 150–170
[29] 刘鑫, 王超, 王沛芳, 等. 营养盐比例对硅藻水华优势种小环藻生长和生理的影响. 环境科学研究, 2021, 34(5): 1196–1204
[30] Guildford S J, Hecky R E. Total nitrogen, total phos-phorus, and nutrient limitation in lakes and oceans: is there a common relationship? Limnology and Oceano-graphy, 2000, 45(6): 1213–1223
[31] 叶玉适, 殷大聪, 陈进, 等. 长江源和怒江源区水体氮磷化学计量特征初探. 长江科学院院报, 2019, 36(9): 6–11
[32] 王丽婧, 田泽斌, 李莹杰, 等. 洞庭湖近 30 年水环境演变态势及影响因素研究. 环境科学研究, 2020, 33(5): 1140–1149
[33] 王岩. 洞庭湖氮磷时空分布及生态风险评价[D]. 哈尔滨: 东北林业大学, 2014
[34] 王艳分, 倪兆奎, 林日彭, 等. 洞庭湖水环境演变特征及关键影响因素识别. 环境科学学报, 2018, 38(7): 2554–2559
Distributions Characteristics of Nutrients and Corresponding Ecological Effects in Dongting Lake
Abstract Based on monitoring data of monthly average COD, TN, and TP concentrations at Chenglingji hydrological station in Dongting Lake from 1990 to 2015, the intra-annual variation, inter-annual variation patterns of nutrients in the water column of Dongting Lake were systematically studied by trend analysis, and further revealed their stoichiometric ratio characteristics and discussed the effects of nutrient concentrations and structure on algal growth. The findings showed that the intra-annual variation patterns of average COD, TN, and TP were similar, indicating that the nutrient content in dry season was higher than that in wet season. However, as for multi-year trend, the annual average COD, TN, and TP in Dongting Lake showed a significant increase since 1990, while C:N and C:P ratios decreased and N:P ratio increased over time. The average C:N:P ratio in the water column of Chenglingji station in Dongting Lake was 13:18:1, and redundancy analysis showed that nutrient concentrations and stoichio-metry ratios in the water would affect algae growth, with C:N, C:P and COD playing a dominant role. During the study period, the overall nitrogen and phosphorus content of the water column in Chenglingji station was generally high and has already met the algae growth demand. In order to avoid adverse ecological consequences such as harmful algal bloom events, external nitrogen and phosphorus inputs should be controlled to maintain the stoichiometric ratio balance in the water column of Dongting Lake.
Key words Dongting Lake; nutrient; stoichiometry; trend analysis