 (1)
(1)doi: 10.13209/j.0479-8023.2023.034
国家自然科学基金(41771049)资助
收稿日期: 2022–07–04;
修回日期: 2022–07–20
北京大学学报(自然科学版) 第59卷 第4期 2023年7月
Acta Scientiarum Naturalium Universitatis Pekinensis, Vol. 59, No. 4 (July 2023)
摘要 为了准确地模拟观赏植物春花期和秋叶期的发生时间, 揭示城市植物物候对气候变化的响应特征并预测赏花和观叶的最佳时机, 利用统一促进模型、统一冷激模型与日最低温–光周期耦合模型, 分别模拟北京植物园 1979—2019 年 20 种春花植物开花期和 10 种秋叶植物叶变色期的年际变化, 并评价最优模型模拟与预测的误差。结果表明, 园林观赏植物春季开花期的发生主要受生态休眠期和生长期促进温度驱动, 内休眠期冷激温度需求没有对春季开花期的发生起到限制作用; 观赏植物秋季叶始变色期的发生主要受温度降低诱发叶片衰老的过程驱动, 叶全变色期的发生主要受光周期缩短诱发叶片衰老的过程驱动。开花始期和开花盛期模拟的平均均方根误差(RMSE)分别为 3.7 和 3.2d, 且这两个物候期的模拟误差具有较好的种间同步性; 叶始变色期和叶全变色期模拟的平均 RMSE 分别为 9.4 和 5.6d, 而这两个物候期之间的模拟误差不具有种间同步性; 物候模型对各种植物开花日期和叶变色日期模拟的 RMSE 与其年际波动标准差呈显著的正相关关系。最优春季和秋季物候过程模型的外推检验与模拟检验的 RMSE 数值相近, 表明模型具有较高的稳健性。
关键词 城市景观时序美; 开花始期和盛期; 叶始变色期和全变色期; 气候驱动; 过程模拟与预测
植物物候现象指以年为周期的植物生长和繁殖现象, 如发芽、展叶、开花、果熟、叶变色和落叶等。这些物候现象的发生时间不仅是环境季节性变化的敏感指示器[1–5]和地表与大气之间二氧化碳、水分和能量交换的调控阀[6–11], 也是自然景观时序美的节律钟。近 30 年来, 大量的物候研究集中揭示了其指示器和调控阀的功能, 对其时序美节律的探索却非常薄弱。随着我国城镇化的快速发展, 城市园林景观设计越来越注重植物形态和色彩的时序配置与空间布局, 而时序配置的关键就是根据各种植物的物候现象在一年之内发生日期的顺序进行园林植物的季相造景, 使人工植物群落在不同的季节都能呈现赏心悦目的造景效果, 形成时序美的动态特征[12]。北京地处暖温带大陆性季风气候区, 四季分明, 植物种类丰富, 且具有明显的季相特征[13–14], 准确地预测具有季节指示意义的观赏植物春花期与秋叶期的发生时间, 可以为园林造景时序美和时令旅游资源的合理开发提供科学依据。
为了预估植物物候现象的发生日期, 需要建立基于过程的物候模型。到目前为止, 物候过程模型的应用集中在对气候变化敏感的指示植物物候期的模拟方面, 很少用来进行城市园林植物观赏物候期的建模。以往的研究表明, 温带落叶树木春季物候现象的发生主要由内休眠期的冷激温度和生态休眠期的促进温度驱动[15–17], 最简单的一阶段过程模型只考虑促进温度对生态休眠期打破和芽生长过程的影响, 较为复杂的两阶段模型则同时考虑冷激温度和促进温度对内休眠期和生态休眠期打破以及芽生长过程的影响。根据芽生长过程对温度的响应形式, 这两类模型又可分为线性的和非线性的两种模拟方式。Chuine[15]在分析不同模型的同型性基础上, 利用生长曲线实现模型的归一化, 分别构建统一促进模型(unified forcing model, UniForc)和统一冷激模型(unified chilling model, UniChill)。将 UniForc和 UniChill 模型应用于我国温带至北热带春季物候过程模拟的研究中, 发现生长在温带的榆树和楝树的展叶和开花期主要受促进温度控制[18–19], 证明Uniforc 模型在温带落叶树种春季物候模拟中具有较高的精确度和稳健性[15,18]。
相比之下, 秋季物候发生的环境归因比较复杂, 过程模型的研究相对滞后。现有的模型包括只考虑秋季低温的寒冷度日模型(cold-degree-day model, CDD)[20]和光周期调节的寒冷度日模型(Delpierre model, DM)[21]。Lang 等[22]提出低温和光周期耦合模型(low temperature and photoperiod multiplicative model, TPM), 综合考虑光周期缩短和日最低温度降低在启动和胁迫叶片衰老过程中的作用。该模型有效地拟合了青藏高原 1981—2012 年 18 个站点 10种木本植物的叶始变色期和 17 种草本植物的黄枯普期的年际变化, 模拟精度明显高于 DM 模型。该模型的普适性已通过对中国北方温带 90 余个站点 6个树种 1981—2014 年叶始变色期和落叶末期的模拟[23]得到初步证实。
Chen 等[24]利用北京植物园 1979—2013 年的木本植物物候观测数据, 揭示出 3 个层次的物候节律, 即顺序相关性节律、准年周期性节律和超年周期性节律, 并检验了它们与温度节律的一致性, 可为利用植物物候发生时间之间的统计关系进行物候预测提供理论支撑。在此基础上, 本文尝试将过程模型引入观赏植物物候的模拟与预测, 利用统一促进模型(UniForc)、统一冷激模型(UniChill)和日最低温和光周期耦合模型(TPM), 分别模拟北京植物园1979—2019 年 20 种春花植物的开花期和 10 种秋叶植物的叶变色期的时间序列, 并评价模拟与预测的精度, 旨在解决以下科学问题: 1)基于本土植物建立的通用物候过程模型能否有效地模拟和预测城市园林观赏植物的开花期和叶变色期; 2)北京市园林观赏植物春季开花期是否主要受到促进温度的驱动, 秋季叶变色期是否主要受到光周期缩短诱发叶片衰老的过程驱动; 3)物候过程模型对观赏植物春季开花期和秋季叶变色期的模拟误差是否存在种间差异性及其可能的原因。
本研究的物候观测站点——北京市植物园(北园)的地理坐标为 40°0′21″N, 116°11′38″E, 位于小西山和寿安山山前洪积扇的顶部, 地势平坦开阔, 背风向阳, 平均海拔约为 60~80m, 因此受阴阳坡和地势等局地小气候条件的影响较小, 可以代表小西山山前一般的自然地理环境条件。该地属暖温带半湿润大陆性季风气候, 四季分明, 夏季炎热多雨, 冬季寒冷干燥, 春季和秋季短暂, 植被的季相变化 明显。
植物物候发生日期的数据来源于个人实地观测记录, 时间为 1979—2019 年。根据《中国物候观测方法》[25], 本研究选取时间序列长度在 15~41 年的20 种春花植物的开花始期(一半以上的观测树木上有一朵或同时有几朵花的花瓣开始完全开放)和开花盛期(观测树木上有一半以上的花蕾展开花瓣), 10 种秋叶植物的叶始变色期(观测树木上的叶子在秋天第一批开始变色)和叶全变色期(观测树木上的所有叶子完全变色)的记录作为研究的物候数据集。按照开花始期的早晚顺序, 春花植物包括蜡梅(Chimonanthus praecox)、迎春花(Jasminum nudiflo-rum)、山桃(Prunus davidiana)、连翘(Forsythia sus-pensa)、玉兰(Yulaniadenudata)、杏(Prunus arme-niaca)、紫玉兰(Yulanialiliiflora)、榆叶梅(Prunus triloba f. multiplex)、贴梗海棠(Chaenomeles spe-ciosa)、紫叶李(Prunus cerasifera f. atropurpurea)、紫丁香(Syringa oblata)、白丁香(Syringa oblata var. alba)、西府海棠(Malus × micromalus)、二色桃(Prunus persica ‘Erse Tao’)、棣棠(Kerria japonica)、毛泡桐(Paulownia tomentosa)、紫藤(Wisteria sinen-sis)、黄刺玫(Rosa xanthina)、太平花(Philadelphus pekinensis)和珍珠梅(Sorbaria sorbifolia), 按照叶始变色期的早晚顺序, 秋叶植物包括黄金树(Catalpa speciosa)、白蜡树(Fraxinus chinensis)、白桦(Betula platyphylla)、一球悬铃木(Platanus occidentalis)、玉兰(Yulaniadenudata)、栾树(Koelreuteria pani-culata)、银杏(Ginkgo biloba)、紫薇(Lagerstroemia indica)、水杉(Metasequoia glyptostroboides)和元宝槭(Acertruncatum)。
气象数据取自北京市海淀气象站, 包括 1978—2019 年的逐日平均气温和逐日最低气温, 该站与北京植物园的直线距离为 6.9km。一般而言, 平原地区气象站代表的范围在 10km 之内, 且海淀气象站的高程(46m)与植物园的平均高程相差不大, 因此利用该站的气象数据进行植物园的物候模拟是可行和现实的。
1.2.1春季物候过程模型
UniForc 模型假设: 在打破内休眠的冷激温度需求满足的条件下, 树木春季物候现象的发生日期取决于越冬芽内休眠解除后促进温度的累积。当促进温度的状态 Sf在第 y 天达到临界值 F*时, 该物候现象发生(式(1))。促进温度的状态 Sf 是按照某一速率的气温累积, 一般将累积气温的起始日期 t1 定为当年的 1 月 1 日, 累积速率 Rf 则是日平均气温 xt 的生长曲线函数(式(2))。UniForc 模型共有 3 个待拟合参数: d, e, F*。
(1)
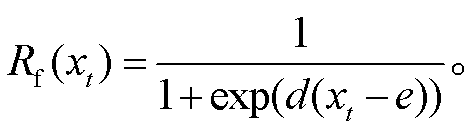 (2)
(2)
UniChill 模型假设: 在冷激温度需求不能满足,并对促进温度累积过程产生影响的条件下, 树木春季物候现象的发生日期由越冬芽内休眠期间的冷激温度和内休眠解除后的促进温度共同决定。当冷激温度的状态 Sc 在第 t1 天达到临界值 C*时, 叶芽或花芽打破内休眠期, 进入生态休眠期(式(3))。冷激温度的状态 Sc 是按照某一速率的气温累积, 一般将累积气温的起始日期 t0 定为前一年的 9 月 1 日, 累积速率则被定义为日平均气温 xt 的生长曲线函数(式(4))。当叶芽或花芽在第 t1 天解除内休眠状态后, 树木开始进入生态休眠期, 并开启促进温度的累积过程, 其累积速率及临界条件同式(2)和(1), 当促进温度的状态 Sf 在第 y 天达到临界值 F*时, 春季物候现象发生。UniChill 模型共有 7 个待拟合参数: a, b, c, C*, d, e, F*。
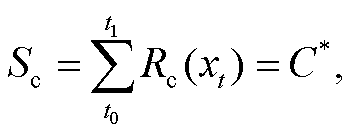 (3)
(3)
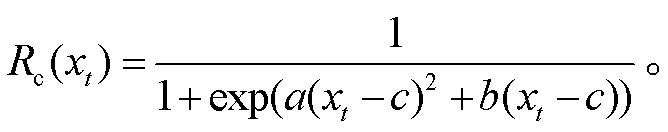 (4)
(4)
1.2.2秋季物候过程模型
TPM 模型假设: 当光周期或日最低温度达到阈值时, 植物叶片开始衰老, 衰老状态由低温和光周期的相互作用控制。随着秋季日最低气温的降低和光周期的缩短, 叶片衰老速率加快, 当叶片衰老状态(Ssen)在第 Y 天达到临界值  时, 秋季物候现象发生(式(5))。叶片衰老状态(Ssen)是从起始日期(Dstart)到秋叶物候发生日的逐日叶片衰老速率(Rsen)之和, 其中 Dstart 可以通过两种方式确定: 如果叶片衰老的开始由光周期缩短触发, Dstart 定义为在光周期最长日期(夏至日, 即一年的第 173 天)之后, 光周期小于光周期阈值(Pstart)的第一天(式(6)), 称为TPMp模型; 如果叶片衰老的开始由温度降低触发, Dstart 就定义为在多年平均日最低气温峰值日期(大致是一年的第 200 天)之后, 日最低气温低于温度阈值(Tstart)的第一天(式(7)), 称为 TPMt 模型。叶片逐日衰老速率(Rsen)是日最低气温和当日光周期算术乘积的生长曲线函数, 它随着该算术乘积的减小而增大(式(8))。TPM 模型共有 4 个待拟合参数: a, b, Pstart/Tstart和。
时, 秋季物候现象发生(式(5))。叶片衰老状态(Ssen)是从起始日期(Dstart)到秋叶物候发生日的逐日叶片衰老速率(Rsen)之和, 其中 Dstart 可以通过两种方式确定: 如果叶片衰老的开始由光周期缩短触发, Dstart 定义为在光周期最长日期(夏至日, 即一年的第 173 天)之后, 光周期小于光周期阈值(Pstart)的第一天(式(6)), 称为TPMp模型; 如果叶片衰老的开始由温度降低触发, Dstart 就定义为在多年平均日最低气温峰值日期(大致是一年的第 200 天)之后, 日最低气温低于温度阈值(Tstart)的第一天(式(7)), 称为 TPMt 模型。叶片逐日衰老速率(Rsen)是日最低气温和当日光周期算术乘积的生长曲线函数, 它随着该算术乘积的减小而增大(式(8))。TPM 模型共有 4 个待拟合参数: a, b, Pstart/Tstart和。
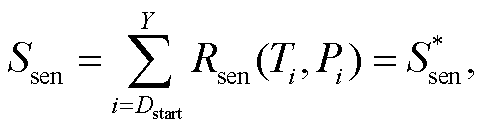 (5)
(5)
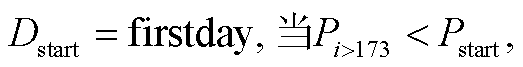 (6)
(6)
 (7)
(7)
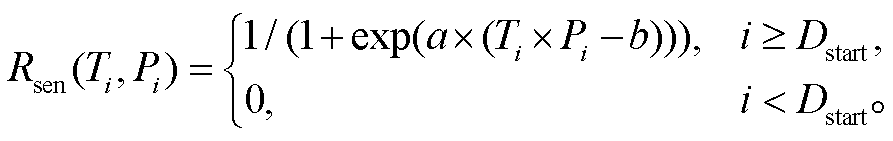 (8)
(8)
1.2.3模型的检验与评价
首先, 根据均方根误差(RMSE, 式(9))最小的原则, 分别确定 UniForc, UniChill, TPMp 和 TPMt 在逐个物种–物候期时间序列模拟中的最优参数组合, 其中模型参数的寻优算法采用模拟退火算法[26]。然后, 筛选各植物各物候期模拟的最优模型, 具体方法是, 比较最优参数组合的 UniForc 和 UniChill模拟的赤池信息量准则(AICc, 式(10))[27], 选择AICc 较小的模型为最优春季物候模型; 比较最优参数组合的 TPMp 和 TPMt 模拟的 RMSE, 选择RMSE 较小的模型作为最优秋季物候模型。在此基础上, 利用纳什效率系数(NSE, 式(11))[28]检验模型模拟的有效性。NSE>0 表示模型有效, 即优于平均值模型, 且 NSE 越接近 1, 模型有效性越高; NSE<0表示模型无效。同时, 利用相关系数(式(12))评价最优模型模拟物候发生日期年际变化的优劣, 模拟与观测时间序列之间的正相关通过 p<0.05 的显著性检验, 表示模型优良; 正相关不显著或负相关, 表示模型较差。
 (9)
(9)
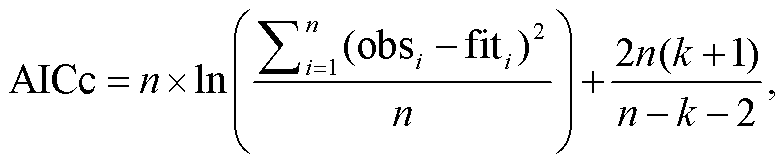 (10)
(10)
 (11)
(11)
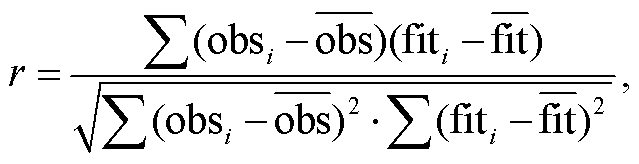 (12)
(12)
式中, obsi 为第 i 年观测日期(序日),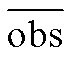 为多年平均观测日期, fiti 为第 i 年模拟日期,
为多年平均观测日期, fiti 为第 i 年模拟日期, 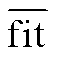 为多年平均模拟日期, k 为模型的参数个数, n 为年数。
为多年平均模拟日期, k 为模型的参数个数, n 为年数。
此外, 为了进一步评价模型外推建模时段以外年份物候发生日期的能力和模型的稳健性, 本文还运用留一交叉验证法[29]对模型进行时间外推检验。该方法适用于小样本数据。具体来说, 首先从原始物候时间序列中选择 n−1 年的数据进行建模, 并获得最优的拟合参数组合; 然后, 利用得到的最优拟合参数外推未参与建模那一年份的物候发生日期, 如此重复 n 次, 使得所有年份的物候发生日期都得到外推检验; 最后, 计算 n 次外推日期与观测日期之间的 RMSE。
在对 20 种植物春季开花日期的模拟中, 开花始期以 UniForc 为最优模型的物种比例达 95%, 以UniChill 为最优模型的只有迎春花 1 种; 开花盛期以 UniForc 为最优模型的物种比例为 80%, 以 UniChill为最优模型的包括杏、白丁香、二色桃和棣棠 4种。最优春季物候过程模型对所有开花始期和开花盛期时间序列的模拟都是有效的(即 NSE>0)。
开花始期的模拟误差(RMSE)平均值为 3.7d, 其中 RMSE 小于 3d 的 5 种植物(占 25%)是玉兰、杏、白丁香、西府海棠和珍珠梅, 有 12 种植物(占60%)的 RMSE 在 3~5d 之间。开花盛期模拟的RMSE 平均值为 3.2d, 其中, 有 9 种植物(占 45%)的RMSE 在 3d 以内, 有 10 种植物(占 50%)的 RMSE 在3~5d 之间(图 1(a))。总的来看, 开花盛期的模拟精度高于开花始期, 其中最为突出的是蜡梅、贴梗海棠、紫叶李、二色桃和棣棠, 它们开花盛期与开花始期模拟的 RMSE 之差都在 1d 以上。此外, 开花始期和开花盛期的模拟误差具有较好的种间同步性, 即如果同一种植物开花始期的模拟误差较小, 则开花盛期的模拟误差也较小; 反之亦然。不同树种两个物候期模拟 RMSE 序列之间的相关系数为0.79 (p<0.001)(图 1(a))。
各种植物开花始期和开花盛期的模拟日期与观测日期之间均呈显著的正相关关系(p<0.001), 表明最优模型能够很好地模拟植物花期的年际波动特征。开花始期的模拟日期与观测日期之间的相关系数平均值为 0.86, 其中有 5 种植物(占 25%)的相关系数大于 0.9, 有 12 种植物(占 60%)的相关系数在0.8~0.9 之间。开花盛期的模拟日期与观测日期之间的相关系数平均值为 0.83, 其中有 4 种植物(占20%)的相关系数大于 0.9, 有 8 种植物(占 40%)的相关系数在 0.8~0.9 之间。从总体上看, 开花始期模拟与观测日期之间的相关系数略大于开花盛期, 差别最显著的是蜡梅、迎春花、紫藤、太平花和珍珠梅。此外, 开花始期和盛期模拟与观测日期之间的相关系数也具有较好的种间同步性, 不同树种两个物候期模拟与观测日期之间相关系数序列的相关系数为0.75 (p<0.001)(图 1(b))。
在对 10 种植物叶始变色期的模拟中, 除黄金树和白桦是以光周期缩短诱发叶片衰老的 TPMp 为最优模型外, 其余 8 种植物均以温度降低诱发叶片衰老的 TPMt 为最优模型; 叶全变色期模拟的最优模型占比情况则相反, 其中白蜡树、黄金树和紫薇是以温度降低诱发叶片衰老的 TPMt 为最优模型, 其余 7种植物是以光周期缩短诱发叶片衰老的 TPMp 为最优模型。最优秋季物候过程模型对所有叶始变色期和叶全变色期模拟的 NSE 均大于 0, 表明模型的模拟具有有效性。
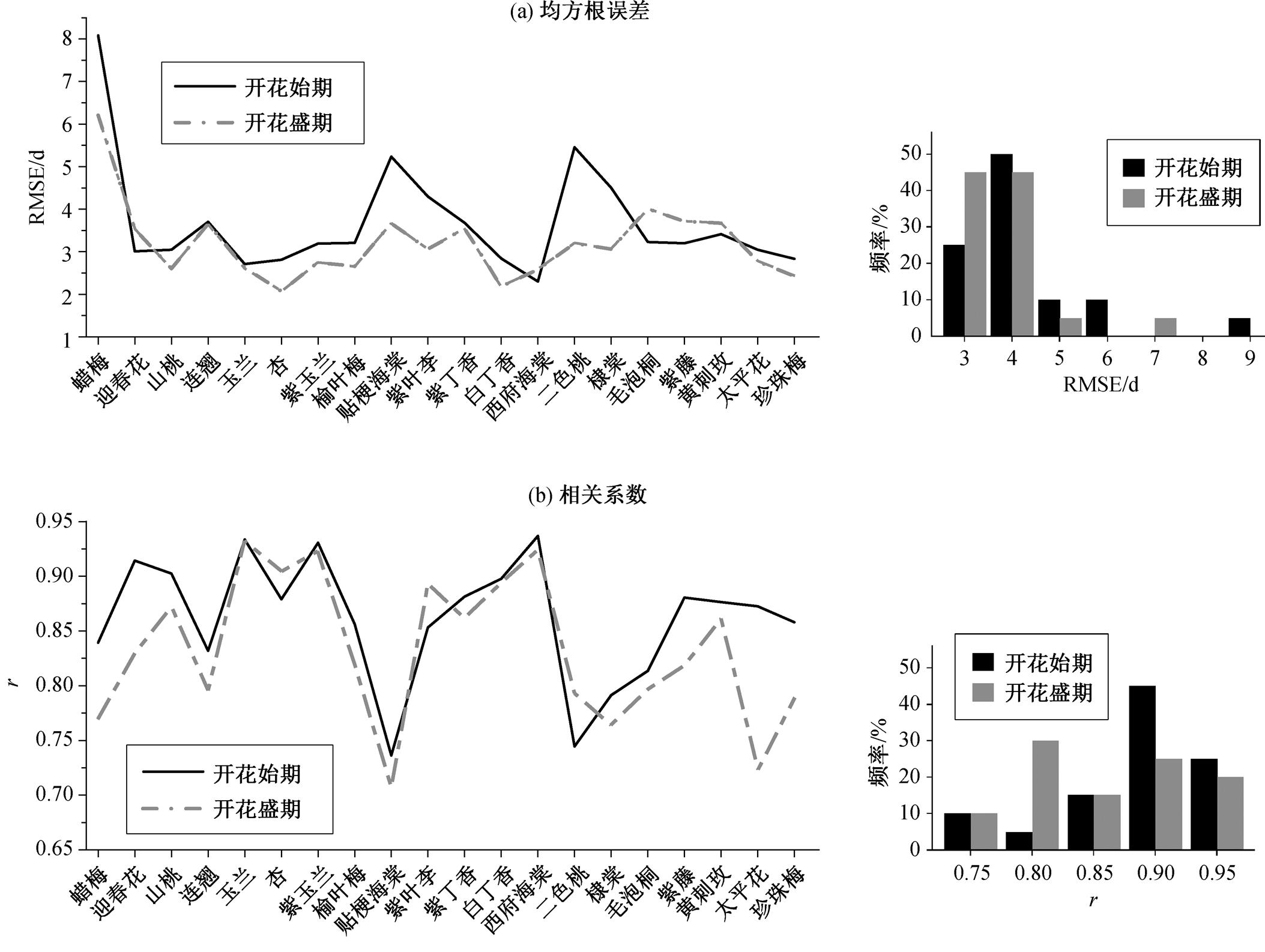
(a)和(b)中的柱状图分别表示RMSE和r的频率分布(下同)
图1 每种植物开花期模拟日期的均方根误差(a)和模拟与观测日期之间的相关系数(b)
Fig. 1 Root mean square error (RMSE) of simulated flowering dates (a) and correlation coefficient (r) between simulated and observed flowering dates (b) for each plant specie
叶始变色期的模拟误差(RMSE)平均值为 9.4d, 其中 RMSE 小于 6d 的只有元宝槭一种植物, RMSE在 6~9d 之间的有 5 种, RMSE大于 9d 的有 4 种。叶全变色期模拟的 RMSE 平均值为 5.6d, 其中 RMSE小于 6d 的有 8 种, RMSE 在 6~9d 之间和大于 9d 的分别只有一种。由此可见, 叶全变色期的模拟误差明显小于叶始变色期, 减小的幅度超过 4d 的是白蜡树、玉兰、栾树和水杉。与开花始期和开花盛期的模拟相比, 叶始变色期和叶全变色期模拟误差的种间同步性明显降低(图 2(a)), 不同树种两个物候期模拟RMSE 序列之间的相关系数仅为 0.22(p>0.05), 未通过显著性检验。
叶始变色期模拟日期与观测日期之间的相关系数平均值为 0.48, 其中有 6 种植物显著正相关(p< 0.05)。叶全变色期模拟日期与观测日期之间均显著正相关(p<0.05), 相关系数平均值为 0.61, 其中相关系数大于 0.6 的有 5 种, 相关系数在 0.4~0.6 之间的有 4种。相比之下, 7 种植物叶全变色期模拟日期与观测日期之间的相关系数大于叶始变色期。此外, 叶始变色期和叶全变色期模拟日期与观测日期之间相关系数的种间同步性也明显低于花期(图2(b)), 不同树种两个物候期模拟日期与观测日期之间相关系数序列的相关系数为 0.34 (p>0.05), 也未通过显著性检验。
利用留一交叉验证法, 我们对每种植物物候期时间序列的最优模型进行模拟年份之外的时间外推检验。结果显示, 外推检验与模拟检验 RMSE 的分布情况较为一致, 外推 RMSE 均大于模拟 RMSE。具体来说, 开花始期和开花盛期的外推 RMSE 平均值分别为 4.3d 和 3.7d, 比开花始期和开花盛期模拟RMSE 的均值分别大 0.6d 和 0.5d (图 3(a)和(b)); 叶始变色期和叶全变色期的外推RMSE平均值分别为11.0d 和 6.7d, 比叶始变色期和叶全变色期模拟RMSE 的均值分别大 1.5d 和 1.2d (图 3(c)和(d))。
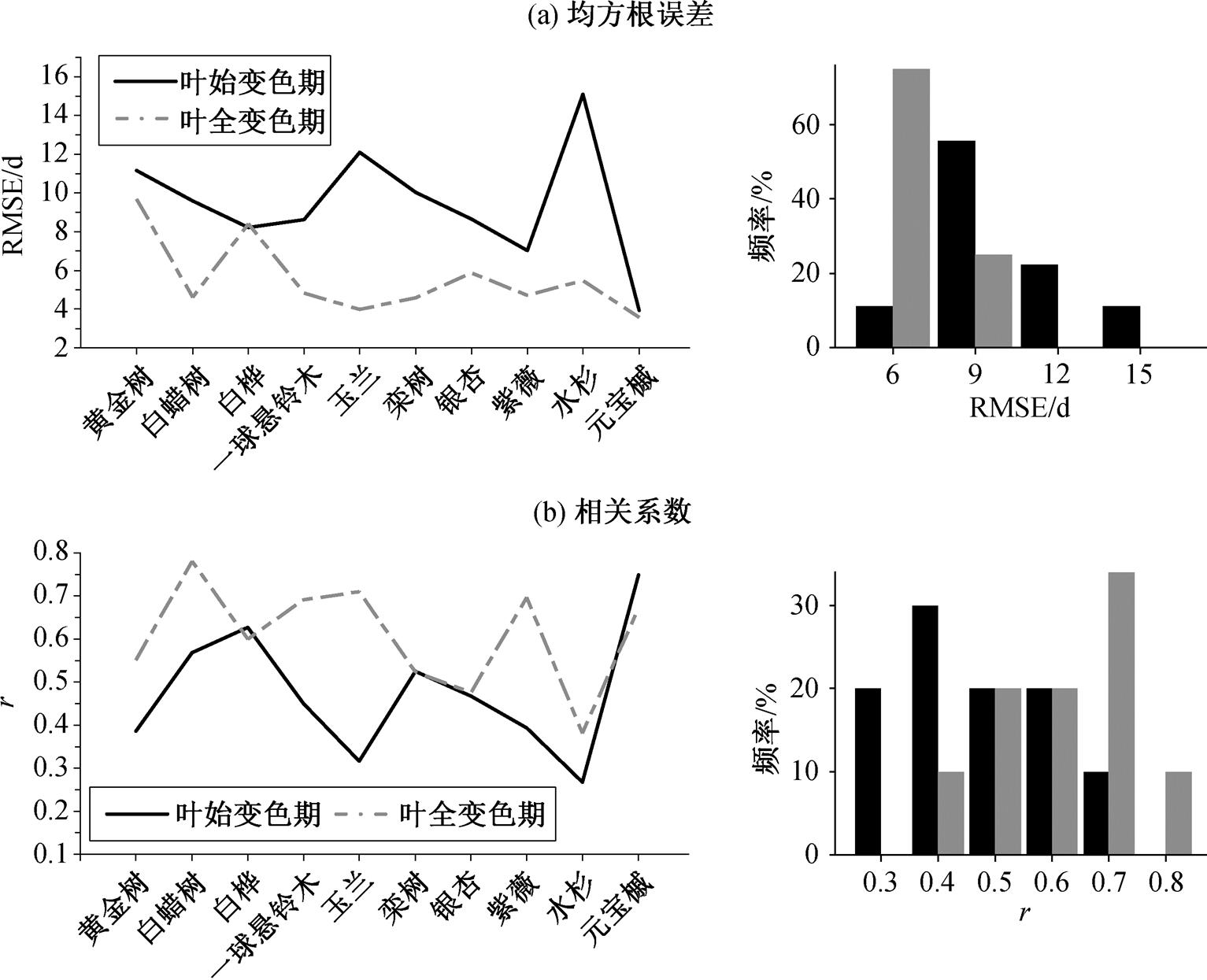
图2 每种植物叶变色期模拟日期的均方根误差(a)和模拟与观测日期之间的相关系数(b)
Fig. 2 RMSE of simulated leaf coloration dates (a) and correlation coefficient between simulated and observed leaf coloration dates (b) for each plant specie

图3 每种植物物候期模拟与外推日期RMSE的分布
Fig. 3 Distribution of root mean square errors (RMSE) of simulated and extrapolated phenophase dates
进一步分析每种植物外推检验与模拟检验误差之间的差别, 可以看出, 各种植物开花始期的外推检验与模拟检验 RMSE 的差值(ΔRMSE)介于 0.2~ 1.2d 之间, 开花盛期的 ΔRMSE 介于 0.1~1.0d 之间(图 4(a)); 各种植物叶始变色期的 ΔRMSE 介于 0.5~ 2.5d 之间, 仅白蜡树和栾树的 ΔRMSE 大于 2d, 叶全变色期的 ΔRMSE 介于 0.4~2.6d 之间, 仅黄金树的 ΔRMSE 大于 2d (图 4(b))。
综上所述, 最优春季和秋季物候过程模型的外推和模拟检验的精度比较接近, 表明本文选用的过程模型在模拟北京植物园典型春花植物花期和秋叶植物叶变色期方面具有较好的稳健性, 为春季和秋季园林观赏植物物候期的预测提供了有效且适用的工具。
本文的研究结果显示, 开花始期和开花盛期以UniForc 为最优模型的物种比例(分别为 95%和 80%)均高于以 UniChill 为最优模型的物种比例, 表明打破内休眠的冷激温度需求在植物响应促进温度之前已经满足, 对春季物候的发生没有限制作用。园林观赏植物春季开花期的早晚主要取决于促进温度, 与中国北方温带地区 136 个站点上 4 个本土植物展叶始期最优模型以 UniForc 为主(83%)的模拟结 果[18]一致, 促进温度对驱动春季物候发生的主导作用也由其他许多物候过程模拟的研究结果证实[19,30–33]。因此, 虽然在全球气候变暖背景下中国北方的冬季气温显著升高, 但植物休眠期间的冷激温度仍然能够满足其越冬芽打破休眠的需求。本文园林观赏植物开花始期和开花盛期模拟的 RMSE 平均值分别为 3.7d和 3.2d, 也与中国北方 4 种本土植物展叶始期模拟的平均误差(3.7d) [18]相当。
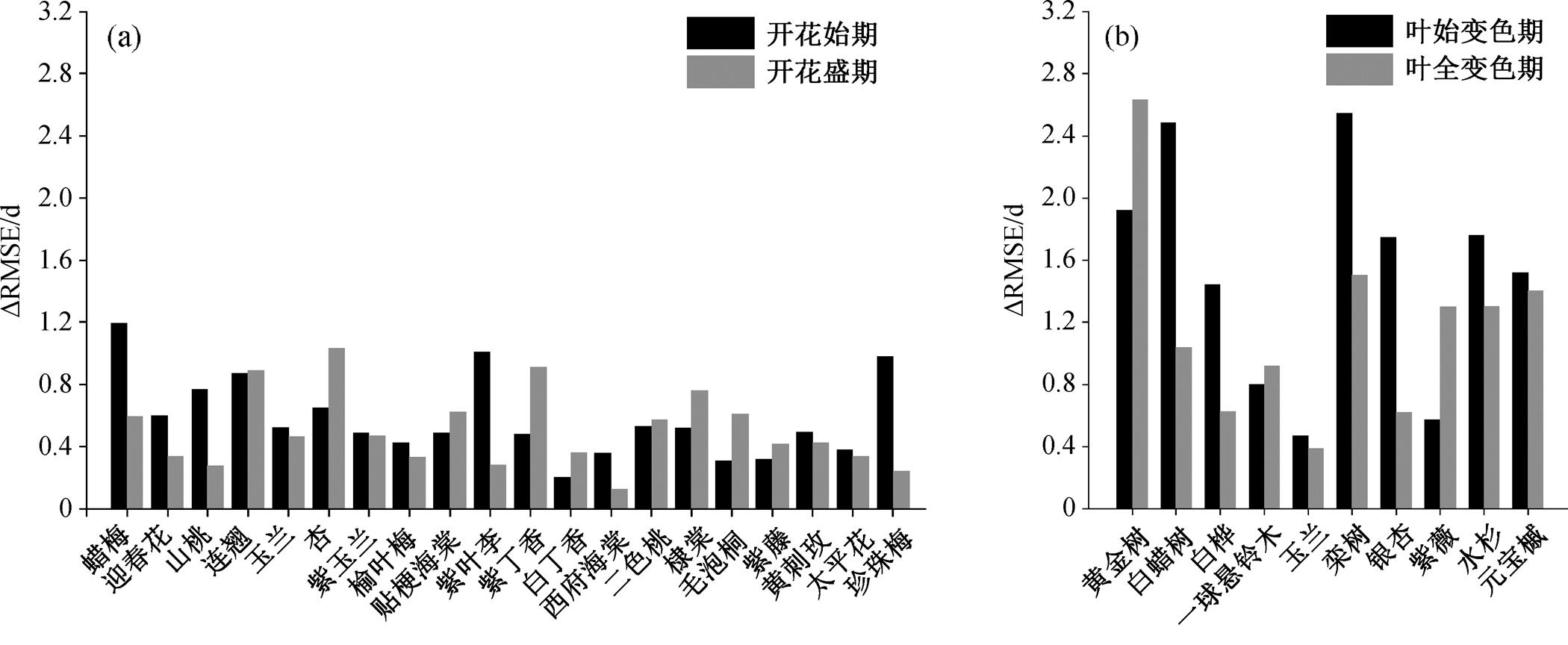
图4 每种植物开花期(a)和叶变色期(b)外推与模拟RMSE之间的差值(ΔRMSE)
Fig. 4 Difference (ΔRMSE) between extrapolated and simulated RMSEs for flowering (a) and leaf coloration (b) of each plant specie
本文对 10 种园林观赏植物叶始变色期的模拟中, 有 8 种植物以温度降低诱发叶片衰老的 TPMt 为最优模型, 两种植物以光周期缩短诱发叶片衰老的TPMp 为最优模型, 与青藏高原 10 个树种 21 个时间序列[22]和中国北方温带地区 6 个树种 185 个时间序列[23]叶始变色期的模拟中主要以 TPMp 为最优模型(>60%)的结果不同。这可能与园林植物秋季物候受气候因子和人为管理措施(如灌溉和施肥等)的双重影响有关, 也使得基于气候因子模拟的平均误差(9.4d)略高于青藏高原(8.2d)和中国北方温带地区(6.9d)。由于秋季的灌溉和施肥很可能导致树木叶变色期推迟[34], 所以综合考虑气候和人为管理因素的影响, 是改进秋季物候模型, 以便提高其对园林观赏植物叶始变色期模拟精度的关键。然而, 对叶全变色期的模拟结果表明, 3 种植物以温度降低诱发叶片衰老的 TPMt 为最优模型, 7 种植物以光周期缩短诱发叶片衰老的 TPMp 为最优模型, 与青藏高原[22]和中国北方温带地区[23]秋季物候的模拟结果一致。最优模型模拟叶全变色期的 RMSE 平均值为5.6d, 明显小于北京植物园(本研究)、青藏高原[22]和中国北方温带地区[23]叶始变色期模拟的 RMSE平均值。
进一步的分析表明, 物候过程模型对某一物候期发生日期的模拟误差(RMSE)与该物候期发生日期的年际波动标准差呈显著的正相关关系, 且模拟误差均小于观测日期的年际波动标准差。由于各种植物开花盛期的标准差(4~10d)一般小于开花始期(5~14d), 而叶全变色期的标准差(5~12d)通常小于叶始变色期(6~15d), 所以物候过程模型对开花盛期和叶全变色期的模拟精度更高(图 5), 这也可以解释各种植物物候发生日期模拟误差种间差异的原因。对于观测日期年际波动较大的植物物候期, 模拟误差较大, 反映过程模型对极端天气事件影响下的极早或极晚物候发生日期的模拟精度有所降低。
此外, 植物开花始期和开花盛期的模拟误差以及模拟日期与观测日期之间的相关系数均具有显著的种间同步性(p<0.001), 主要是由于同种植物开花始期和开花盛期的年际变化具有同步提前或推迟的特点, 并且对促进温度的响应过程一致。相比之下, 植物叶始变色期和叶全变色期的模拟误差以及模拟与观测日期之间的相关系数均不具有显著的种间同步性, 主要是由于同种植物叶始变色期和叶全变色期的年际变化不具有同步提前或推迟的特点, 并且这两个物候期的发生对诱发叶片衰老途径(温度降低诱发或光周期缩短诱发两种途径)的选择也不一致。在全球气候增暖的背景下, 极端高、低温和干旱事件频发, 可能引起开花始期与开花盛期对温度的响应过程和叶始变色期与叶全变色期对光周期与温度耦合作用的响应过程发生显著的改变, 从而导致自然景观时序美节律钟时间的调整, 这将增加利用这些模型对春花和秋叶物候观赏最佳时间预测的不确定性。

图5 植物物候观测日期标准差与模拟误差(RMSE)之间的相关回归分析
Fig. 5 Correlation and regression analyses between standard deviations of observed date of plant phenology and simulated error (RMSE)
本文利用春季和秋季物候过程模型, 分别拟合北京植物园 1979—2019 年 20 种春花植物的开花期和10 种秋叶植物的叶变色期时间序列, 选出每种植物物候时间序列的最优模型, 并对模拟和外推效果进行检验, 得到如下主要结论。
1)基于通用的春季和秋季物候过程模型, 可以有效地模拟和预测城市园林观赏植物的开花期和叶变色期, 其中, 对开花盛期的模拟精度高于开花始期, 对叶全变色期的模拟精度高于叶始变色期。除叶始变色期外, 模型对其他 3 个物候期的模拟精度与对本土植物春季和秋季物候期的模拟精度相当。
2)园林观赏植物与本土植物春季物候对气温变化的响应基本上一致, 表现为大多数开花始期和盛期时间序列的模拟以促进温度驱动的 UniForc 为最优模型。园林观赏植物与本土植物叶变色期对气温和光周期耦合变化的响应不完全相同, 表现为大多数植物的叶始变色期以温度降低诱发叶片衰老的TPMt 为最优模型, 而大多数植物的叶全变色期以光周期缩短诱发叶片衰老的 TPMp 为最优模型。
3)未来气候增暖引起的极端天气事件频发, 很可能显著地影响园林观赏植物物候的时序美节律, 将增加利用本文建立的模型对春花期和秋叶期最佳观赏时间预测的不确定性。
参考文献
[1] Menzel A, Fabian P. Growing season extended in Europe. Nature, 1999, 397: 659–659
[2] Chmielewski F M, Rötzer T. Response of tree pheno-logy to climate change across Europe. Agricultural and Forest Meteorology, 2001, 108(2): 101–112
[3] Menzel A. Plant phenological anomalies in Germany and their relation to air temperature and NAO. Climatic Change, 2003, 57(3): 243–263
[4] Root T L, Price J T, Hall K R, et al. Fingerprints of global warming on wild animals and plants. Nature, 2003, 421: 57–60
[5] 陈效逑, 喻蓉. 1982—1999 年我国东部暖温带植被生长季节的时空变化. 地理学报, 2007(1): 41–51
[6] Myneni R B, Keeling C D, Tucker C J, et al. Increased plant growth in the northern high latitudes from 1981 to 1991. Nature, 1997, 386: 698–702
[7] Black T A, Chen W J, Barr A G, et al. Increased carbon sequestration by a boreal deciduous forest in years with a warm spring. Geophysical Research Letters, 2000, 27(9): 1271–1274
[8] Barr A G, Black T A, Hogg E H, et al. Interannual variability in the leaf area index of a boreal aspen-hazelnut forest in relation to net ecosystem production. Agricultural and Forest Meteorology, 2004, 126(3/4): 237–255
[9] Kljun N, Black T A, Griffis T J, et al. Response of net ecosystem productivity of three boreal forest stands to drought. Ecosystems, 2006, 9(7): 1128–1144
[10] Delpierre N, Soudani K, Francois C, et al. Exceptional carbon uptake in European forests during the warm spring of 2007: a data–model analysis. Global Change Biology, 2009, 15(6): 1455–1474
[11] Chen Xiaoqiu, Xu Lin. Phenological responses of Ulmus pumila (Siberian Elm) to climate change in the temperate zone of China. International Journal of Biometeorology, 2012, 56(4): 695–706
[12] 杨国栋, 陈效逑. 木本植物物候相组合分类研 究——以北京市植物园栽培树种为例. 林业科学, 2000, 36(2): 39–46
[13] 陈效逑, 曹志萍. 植物物候期的频率分布型及其在季节划分中的应用. 地理科学, 1999(1): 22–28
[14] 邢小艺, 郝培尧, 李冠衡, 等. 北京植物物候的季节动态特征——以北京植物园为例. 植物生态学报, 2018, 42(9): 906–916
[15] Chuine I. A unified model for budburst of trees. Journal of Theoretical Biology, 2000, 207(3): 337–347
[16] Chuine I, Kramer K, Hänninen H. Phenology: an in-tegrative environmental science. Dordrecht: Kluwer Academic Publishers, 2003
[17] Hänninen H, Kramer K. A framework for modelling the annual cycle of trees in boreal and temperate regions. Silva Fennica, 2007, 41(1): 167–205
[18] Xu Lin, Chen Xiaoqiu. Regional unified model-based leaf unfolding prediction from 1960 to 2009 across northern China. Global Change Biology, 2013, 19(4): 1275–1284
[19] Chen Xiaoqiu, Wang Lingxiao, Inouye D. Delayed response of spring phenology to global warming in subtropics and tropics. Agricultural and Forest Meteo-rology, 2017, 234: 222–235
[20] Dufrêne E, Davi H, Francois C, et al. Modelling carbon and water cycles in a beech forest: Part I: model description and uncertainty analysis on modelled NEE. Ecological Modelling, 2005, 185(2/3/4): 407–436
[21] Delpierre N, Dufrêne E, Soudani K, et al. Modelling interannual and spatial variability of leaf senescence for three deciduous tree species in France. Agricultural and Forest Meteorology, 2009, 149(6/7): 938–948
[22] Lang Weiguang, Chen Xiaoqiu, Qian Siwei, et al. A new process-based model for predicting autumn phe-nology: How is leaf senescence controlled by pho-toperiod and temperature coupling?. Agricultural and Forest Meteorology, 2019, 268: 124–135
[23] 陈奕竹, 郎伟光, 陈效逑. 中国北方树木秋季物 候的过程模拟及其区域差异归因. 植物生态学报, 2022, 46(7): 753–765
[24] Chen Xiaoqiu, Zhang Weiqi, Ren Shilong, et al. Tem-poral coherence of phenological and climatic rhyth-mmicity in Beijing. International Journal of Biome-teorology, 2017, 61(10): 1733–1748
[25] 宛敏渭, 刘秀珍. 中国物候观测方法. 北京: 科学出版社, 1979
[26] Chuine I, Cour P, Rousseau D D. Fitting models pre-dicting dates of flowering of temperate-zone trees using simulated annealing. Plant, Cell & Environment, 1998, 21(5): 455–466
[27] Hurvich C M, Tsai C L. Regression and time-series model selection in small samples. Biometrika, 1989, 76(2): 297–307
[28] Nash J E, Sutcliffe J V. River flow forecasting through conceptual models part I — a discussion of principles. Journal of Hydrology, 1970, 10(3): 282–290
[29] Archetti M, Richardson A D, O’Keefe J, et al. Predic-ting climate change impacts on the amount and dura-tion of autumn colors in a New England forest. PLOS ONE, 2013, 8(3): e57373
[30] Linkosalo T, Lappalainen H K, Hari P. A comparison of phenologicalmodels of leaf bud burst and flowering of boreal trees using independentobservations. Tree Physiology, 2008, 28(12): 1873–1882
[31] Richardson A D, O’Keefe J. Phenological differences between understory and overstory // Phenology of eco-system processes. New York: Springer, 2009: 87–117
[32] Parker A K, De Cortazar-Atauri I G, Van Leeuwen C, et al. General phenological model to characterise the timing of flowering and veraison of Vitis vinifera L. Australian Journal of Grape and Wine Research, 2011, 17(2): 206–216
[33] Fu Yongshuo, Campioli M, Oijen M V, et al. Baye- sian comparison of six different temperature-based budburst models for four temperate tree species. Ecological Modelling, 2012, 230: 92–100
[34] Estiarte M, Peñuelas J. Alteration of the phenology of leaf senescence and fall in winter deciduous species by climate change: effects on nutrient proficiency. Global Change Biology, 2015, 21(3): 1005–1017
Process-Based Simulation of Spring Flowering and Autumn Leaf Coloration of Main Ornamental Plants in Beijing Botanical Garden
Abstract Accurately simulating occurrence dates of spring flowering and autumn leaf coloration of ornamental plants is of significant importance for revealing phenological response of urban vegetation to climate change and predicting the optimum timing for flower and foliage viewing. This study employed the Unified Forcing Model (UniForc) and Unified Chilling Model (UniChill) to fit the first flowering and 50% flowering dates of 20 plant species from 1979 to 2019 in Beijing Botanical Garden, and the Low Temperature and Photoperiod Multiplicative Model (TPM) to fit the first leaf coloration and 100% coloration dates of 10 plant species. The errors of optimum models in simulation and prediction were evaluated. Results show that spring flowering of ornamental plants is mainly driven by forcing temperature during ecodormancy and growth periods, but less restricted by chilling temperature during endodormancy period. First leaf coloration of ornamental plants is mainly driven by the process of leaf senescence induced by daily minimum temperature decrease, and 100% leaf coloration is mainly driven by the process of leaf senescence induced by photoperiod shortening. The average simulated root-mean square errors (RMSE) for first flowering and 50% flowering are 3.7 days and 3.2 days, respectively, and simulated RMSEs for the two spring phenophases show good interspecific synchronization. The average simulated RMSEs for first leaf coloration and 100% leaf coloration are 9.4 days and 5.6 days, respectively, but simulated RMSEs for the two autumn phenophases do not display interspecific synchronization. Simulated RMSEs of flowering dates and leaf coloration dates of various plants correlate significantly and positively with their standard deviation of interannual variations. The simulated and extrapolating RMSEs of optimum spring and autumn process-based models are very close, indicating that the models have high robustness.
Key words timing sequential beauty of urban landscape; first and 50% flowering; first and 100% leaf coloration; climate driving; process-based simulation and prediction