表1 毛乌素沙地樟子松人工林样地概况
Table 1 Basic information of P. sylvestris plantation sample plot in the Mu Us Desert
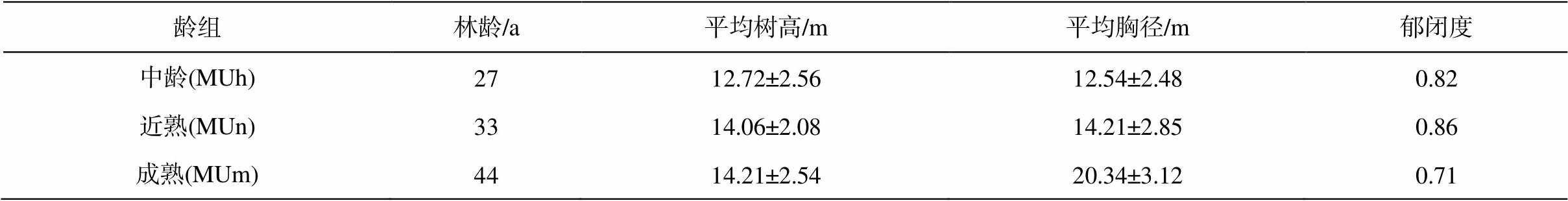
龄组林龄/a平均树高/m平均胸径/m郁闭度 中龄(MUh)2712.72±2.5612.54±2.480.82 近熟(MUn)3314.06±2.0814.21±2.850.86 成熟(MUm)4414.21±2.5420.34±3.120.71
北京大学学报(自然科学版) 第59卷 第3期 2023年5月
Acta Scientiarum Naturalium Universitatis Pekinensis, Vol. 59, No. 3 (May 2023)
doi: 10.13209/j.0479-8023.2023.031
内蒙古自治区科技计划项目(2022YFHH0131)和中央高校基本科研业务费项目(2021ZY47)资助
收稿日期: 2022–05–06;
修回日期: 2022–07–14
摘要 为揭示樟子松人工林根内真菌群落相互关系的动态特征, 以毛乌素沙地樟子松人工林为研究对象, 采用生态网络分析法, 分析不同林龄(中龄、近熟和成熟)及其年内生长阶段(生长季初期、旺盛期和末期)樟子松根内真菌群落共现网络动态特征及其对气候因子的响应, 得到如下结果。1)在不同林龄阶段, 樟子松人工林根内真菌共现网络差异不明显; 在不同生长季阶段, 根内真菌共现网络差异明显, 生长季旺盛期, 根内真菌共现网络较为复杂。2)在不同林龄和不同生长季阶段, 樟子松人工林根内真菌关键类群差异明显, 主要包括地孔菌属(Geopora)、棉革菌属(Tomentella)、暗球腔菌属(Phaeosphaeria)、Neocucurbitaria和被孢霉属(Mortierella)等。生长季初期和末期关键类群为外生菌根真菌和腐生真菌, 旺盛期为腐生真菌和病原真菌。中龄林关键类群为腐生真菌和病原真菌, 成熟林为外生菌根真菌和腐生真菌。3)外生菌根真菌网络的主要影响因子为相对湿度(P<0.05), 腐生真菌和病原真菌网络的主要影响因子为平均降水量和相对湿度。因此, 毛乌素沙地樟子松人工林根内真菌网络在不同生长季阶段的动态变化强于不同林龄阶段, 且生长季旺盛期共现网络的结构较复杂; 根内真菌网络主要受平均降水量和相对湿度影响。腐生真菌对维持不同林龄和不同生长季阶段真菌网络的稳定性均有重要作用, 在较大降水量和湿度的生长季旺盛期, 关键类群为腐生真菌和病原真菌, 对樟子松的生长和健康状况有着潜在的影响。
关键词 沙生植物; 林龄; 生长季; 土壤真菌; 共现网络; 气候因子
根内真菌广泛定殖于植物根系中, 直接或间接地影响着植物对水分和养分的吸收[1], 与寄主植物生长和健康状况密切相关。其中, 外生菌根真菌通过寄主的光合作用获得并固定碳, 作为交换, 为寄主提供磷、氮、钾等矿物质营养和生长素等代谢产物, 增强植物抗逆性[2]; 病原真菌通过感染植物组织抑制其生长繁殖, 诱发植物病害, 造成寄主植物死亡[3]; 腐生真菌则可以通过分泌胞外酶, 分解利用有机质, 抑制病原真菌生长[4]。长期以来, 对根内真菌群落结构的研究多集中于物种丰富度和多样性等方面[5–6], 研究成果对全面认识理解根内真菌群落结构, 服务解析根内真菌生态功能具有重要意义[7]。但是, 根内真菌群落结构复杂, 不同真菌之间存在积极的合作关系、消极的竞争关系或中性相互作用[8]。根内真菌的相互作用对生态系统功能的影响可能比物种丰富度和多样性更重要, 且这种相互作用存在较强的环境敏感性和异质性[9]。
在森林生态系统中, 林木生长和气候与根内真菌相互作用存在密切联系。在不同林龄阶段, 林木有着不同的养分需求, 可直接或间接地对根内真菌群落进行招募, 进而选择性地调节根内真菌之间的相互作用, 塑造与之相适应的根内真菌群落网络[10]。在不同生长季阶段, 林木生长主要受温度和降水的调控[11]。一般来说, 温度变化导致微生物类群的选择性生长, 改变根内真菌之间的相互作用[12]; 降水则直接改变土壤含水量, 根内真菌对土壤水势的耐受性或敏感度也不同, 会影响菌丝获取水分的能力, 也为其相互作用创造了更多的途径和机会[13–14]。因此, 研究林木生长和气候因子对根内真菌互作的影响, 有助于深入理解气候变化背景下林木根内真菌网络的动态特征和生态功能。
长期以来, 测试和分析手段的局限导致对根内真菌相互作用的认识和理解较为薄弱。表征和分析土壤微生物相互作用方法的不断更新, 为认识土壤与微生物的相互关系提供了有力的武器[15]。其中, 生态网络分析法以随机矩阵理论为基础, 具有较高的可靠性和敏感性, 可用于高通量数据的模块化网络构建和功能预测[5], 目前, 已在复杂的森林生态系统中得到广泛应用, 成为揭示土壤微生物在微生境中的共现关系及主要影响因素的重要途经, 对深入认识和理解根内真菌群落的生态功能和稳定性有重要意义[16]。
樟子松(Pinus sylvestris var. mongolica)耐寒, 耐旱, 耐贫瘠, 是我国北方干旱半干旱地区防沙治沙和水土保持工作的重要绿化造林树种[17], 也是典型的菌根依赖型树种, 其生长发育受根内真菌群落复杂相互作用的影响[18]。鉴于此, 本文以毛乌素沙地樟子松人工林为研究对象, 采用生态网络分析法, 研究不同林龄及其年内生长阶段, 樟子松人工林根内真菌群落共现网络的动态特征及其对气候因子的响应, 以期深入理解樟子松人工林根内真菌群落的相互关系, 并为樟子松人工林可持续经营管理提供理论依据。
研究区位于陕西省榆林市红石峡沙地植物园, 地处毛乌素沙地东南部(38°26′N, 109°12′E), 海拔1080m。该地处于半干旱区, 具有暖温带大陆性季风气候特点, 干燥多风, 冬春季节风沙较大, 降水量年际变化率大。年均温度为 9.1°C, 年均日照时数为 2914 小时, 年均降水量为 365.7mm, 年均蒸发量为 2502mm。土壤类型为风沙土, 通透性好, 保水性差, 较贫瘠, 表土结构疏松, 有机质含量低。优势植物种主要有樟子松、蒙古冰草(Agropyron mongolicum)、紫穗槐(Amorpha fruticosa)和花棒(Hedysarum scoparium)等[19]。
2018 年, 选取立地条件和管理措施相同的中龄林(27a)、近熟林(33a)和成熟林(44a)樟子松人工林作为试验样地, 分别随机布设 50m×50m 样方, 每木检尺后分别在各样方中选取 5 株间距至少为 10m的健康标准木(表 1)。在樟子松生长季初期(4—6月)、旺盛期(7—8 月)和末期(9 月)分别采集样品(表2), 铲去树木基部半径 1~2m 内的地表凋落物后, 沿树干根部挖至根的末级, 剔除杂草和其他植物根系, 采集直径小于 2mm 的新鲜细根样品。每株标准木随机采集 3 个重复样本, 并将其充分混合为1 个样品。细根样品配以少量土壤保鲜, 装入塑封袋, 保存于–4°C 移动冰箱中, 带回实验室后–80°C冷冻保存[20]。
表1 毛乌素沙地樟子松人工林样地概况
Table 1 Basic information of P. sylvestris plantation sample plot in the Mu Us Desert
龄组林龄/a平均树高/m平均胸径/m郁闭度 中龄(MUh)2712.72±2.5612.54±2.480.82 近熟(MUn)3314.06±2.0814.21±2.850.86 成熟(MUm)4414.21±2.5420.34±3.120.71
说明: MUh, MUn 和 Mum 分别表示中龄林、近熟林和成熟林, 下同。
表2 毛乌素沙地樟子松人工林气候
Table 2 Climate of of P. sylvestris plantation in the Mu Us Desert
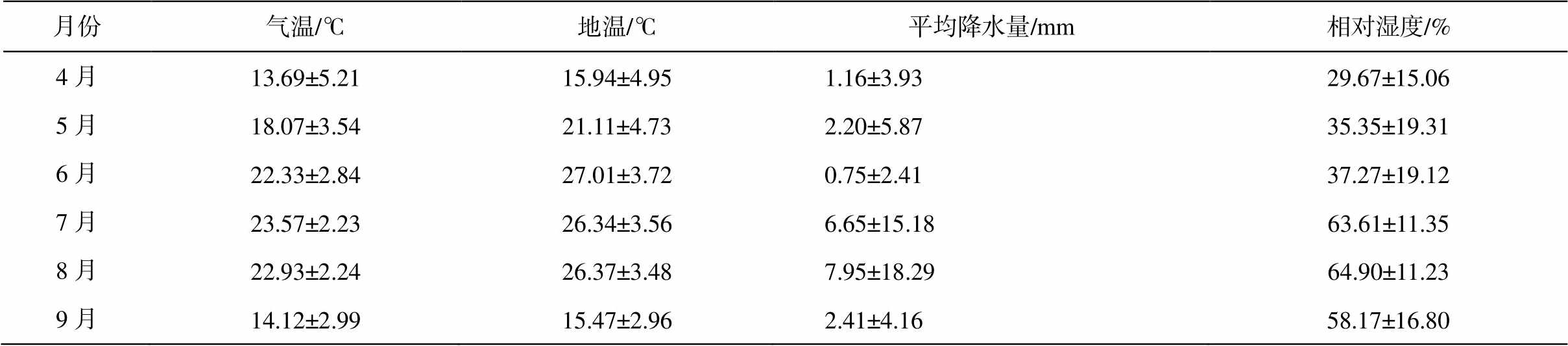
月份气温/℃地温/℃平均降水量/mm相对湿度/% 4月13.69±5.2115.94±4.951.16±3.9329.67±15.06 5月18.07±3.5421.11±4.732.20±5.8735.35±19.31 6月22.33±2.8427.01±3.720.75±2.4137.27±19.12 7月23.57±2.2326.34±3.566.65±15.1863.61±11.35 8月22.93±2.2426.37±3.487.95±18.2964.90±11.23 9月14.12±2.9915.47±2.962.41±4.1658.17±16.80
取出细根样品, 并用蒸馏水小心清洗其表面土壤颗粒和杂物。清洗干净后, 将细根剪成 10cm 的根段, 置于盛有蒸馏水的培养皿中。将根尖端末级根剪下后, 加入液氮, 在研钵中研磨破碎 DNA, 使用 PowerSoil® DNA 试剂盒(MO BIO公司)提取样品中的 DNA, 利用 1%琼脂糖凝胶电泳检测抽提的基因组 DNA。采用 PCR 对根内真菌的 rDNA ITS 区进行扩增, 真菌通用引物为 ITS1F (5′-CTTGGTCATT TAGAGGAAGTAA-3′)和 ITS2 (5′-GCTGCGTTCTT CATCGATGC-3′), 每个样本重复 3 次。PCR 扩增试验采用 25μL 的反应体系: KAPA 2G Robust HotStart ReadyMix 12.5μL, 引物 ITS1 (5μmol/L) 1μL, 引物ITS2 (5μmol/L) 1μL, 模板 DNA 5μL (30ng), ddH2O补足至 25μL。PCR 反应条件如下: 95°C 5min; 95°C 45s, 55°C 50s, 72°C 45s, 28 个循环; 72°C 10min。混合 PCR 产物后, 采用琼脂糖(质量分数 2%)凝胶电泳检测, 选用 AxyPrepDNA 凝胶试剂盒切胶回收PCR 产物, 采用 Tris-HCl 缓冲液(0.05mol/L, 25°C)洗脱后, 用琼脂糖(质量分数 2%)电泳检测。检测合格的 PCR 产物用于 MiSeq 上机测序。测序工作由北京奥维森生物科技有限公司协助完成。
基于 NCBI 数据库, 采用 BLAST 方法, 对比分析 I 代表序列。如果对比相似率≥97%, 则可鉴定到种水平, 如果相似率为 90%~97%, 则可鉴定到属水平。通过 FUNGuild 平台(http://www.stbates.org/gui-lds/app.php), 将樟子松人工林根内真菌划分为不同的营养型和功能群。结果数据仅保留置信度为“很可能” (Probable)和“极可能” (Highly probable)的OTUs 及其类别。
利用高通量测序获得的 OTUs 数据, 对不同林龄(中龄、近熟和成熟)及其年内生长阶段(生长季初期、旺盛期和末期)分别整合数据, 进行网络分析。采用 Pearson 相关性分析法, 分析各 OTU 间的相关性, 筛选并保留 P<0.05 的 OTU, 并采用 Gephi 9.0 软件进行网络构建, 获取网络拓扑特征参数。使用 Maslov-Sneppen 方法进行随机网络构建, 在不改变原有网络节点和连线数的基础上, 重新连接原网络中不同位置的节点, 并构建 1000 次随机网络, 比较分子生态网络与随机网络之间的差别。根据节点的模块内连通性(Zi)和模块间连通性(Pi), 将网络中所有节点的模块角色分为 4 种类型, 分别是模块枢纽(module hubs)、连接器(connectors)、网络枢纽(network hubs)以及外围节点(peripherals)。Zi 和 Pi表征节点作用, 一般将 Zi≥2.5 或 Pi≥0.62 的节点定为关键物种。因此, 落在模块枢纽、网络枢纽和连接器的节点在模块间及各自模块内起重要作用。
气象数据来自中国气象数据网。使用 R 4.0.3进行 Mantel 分析。使用 SPSS 26, 通过单因素方差分析法, 分析真菌群落多样性指数以及网络结构差异的显著性。使用 Origin 2018 软件和 R 4.0.3 绘图, Gephi 9.0和 Cytoscape 3.9.0 软件对网络进行可视化处理。
如表 3 所示, 毛乌素沙地樟子松人工林根内真菌群落经验网络模块化指数、平均路径长和平均聚类系数均大于随机网络的相应数值, 符合网络特征。在不同林龄阶段, 共现网络差异不明显; 在不同生长季阶段, 根内真菌群落共现网络存在明显差异(表 3, 图 1 和 2)。生长季旺盛期节点数和边数明显高于生长季初期和末期, 平均路径长度最小, 但平均度最高。除生长季初期外, 各 OTU 之间均出现负相关, 但各林龄和生长季呈正相关的比例均高于呈负相关的比例。此外, 不同林龄阶段, 外生菌根真菌、腐生真菌和病原真菌的数量变化不明显, 在不同生长季阶段, 3 类功能群数量均先增大后减小, 生长季旺盛期达到最大值。
如图 3 所示, 毛乌素沙地不同林龄樟子松根内真菌群落网络节点落于连接器和模块枢纽区域, 近熟林和成熟林模块枢纽节点分别为 2 和 5 个 OTU, 中龄林均为连接器节点, 无模块枢纽节点。不同生长季均无模块枢纽和网络枢纽节点, 起重要作用的节点均落于连接器区域。生长季内各月连接器节点OTU 分别为 14, 21, 35, 74, 38 和 38 个, 生长季旺盛期位于连接器区域的 OTU 数量和属数量达到最高值(图 4)。
表3 毛乌素沙地樟子松人工林根内真菌群落的网络拓扑特征
Table 3 Topological characteristics of fungal community network in roots of P. sylvestris plantation in the Mu Us Desert
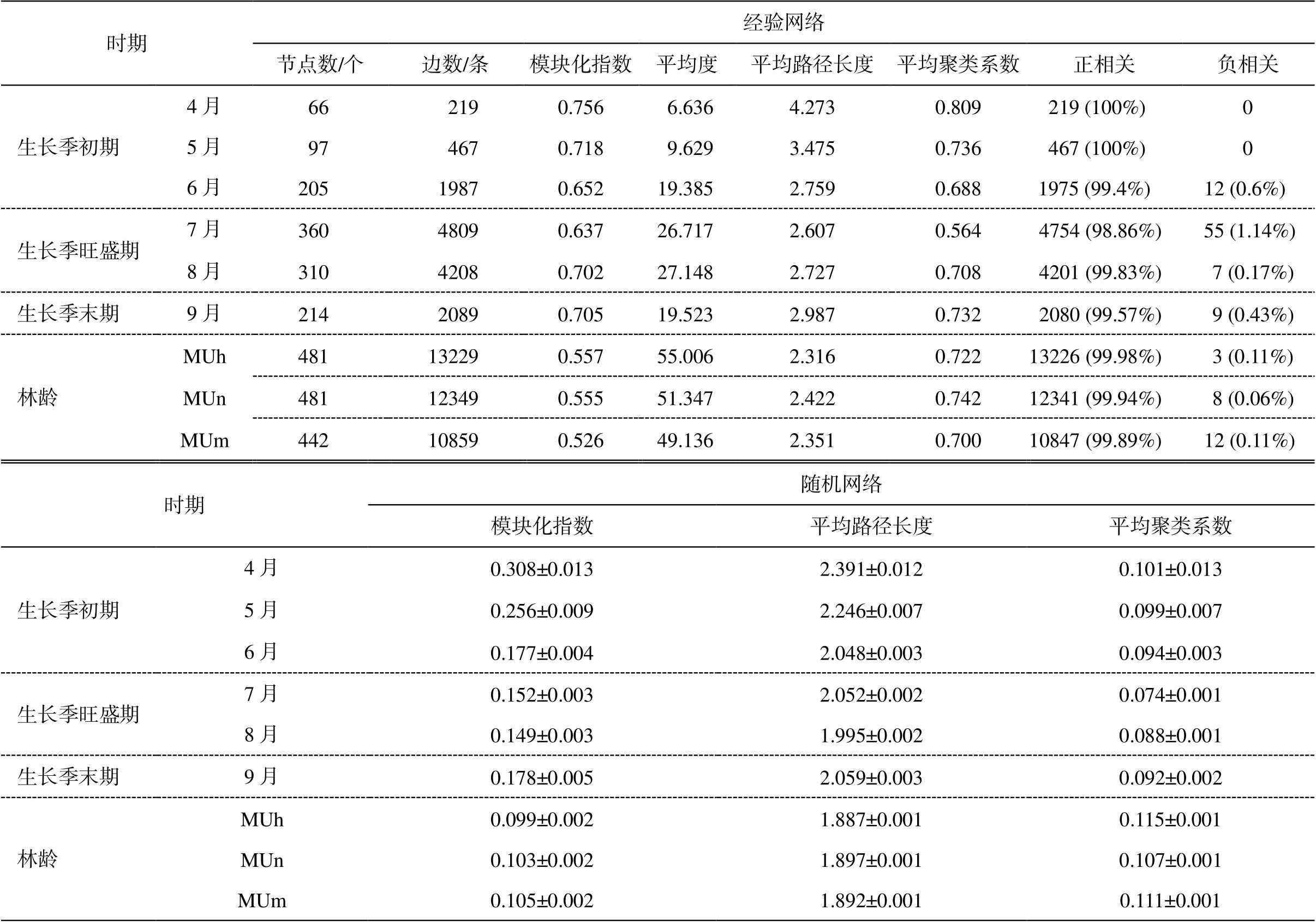
时期经验网络 节点数/个边数/条模块化指数平均度平均路径长度平均聚类系数正相关负相关 生长季初期4月662190.7566.6364.2730.809219 (100%) 0 5月974670.7189.6293.4750.736467 (100%) 0 6月20519870.65219.3852.7590.6881975 (99.4%) 12 (0.6%) 生长季旺盛期7月36048090.63726.7172.6070.5644754 (98.86%)55 (1.14%) 8月31042080.70227.1482.7270.7084201 (99.83%)7 (0.17%) 生长季末期9月21420890.70519.5232.9870.7322080 (99.57%)9 (0.43%) 林龄MUh481132290.55755.0062.3160.72213226 (99.98%)3 (0.11%) MUn481123490.55551.3472.4220.74212341 (99.94%)8 (0.06%) MUm442108590.52649.1362.3510.70010847 (99.89%)12 (0.11%) 时期随机网络 模块化指数平均路径长度平均聚类系数 生长季初期4月0.308±0.0132.391±0.0120.101±0.013 5月0.256±0.0092.246±0.0070.099±0.007 6月0.177±0.0042.048±0.0030.094±0.003 生长季旺盛期7月0.152±0.0032.052±0.0020.074±0.001 8月0.149±0.0031.995±0.0020.088±0.001 生长季末期9月0.178±0.0052.059±0.0030.092±0.002 林龄MUh0.099±0.0021.887±0.0010.115±0.001 MUn0.103±0.0021.897±0.0010.107±0.001 MUm0.105±0.0021.892±0.0010.111±0.001
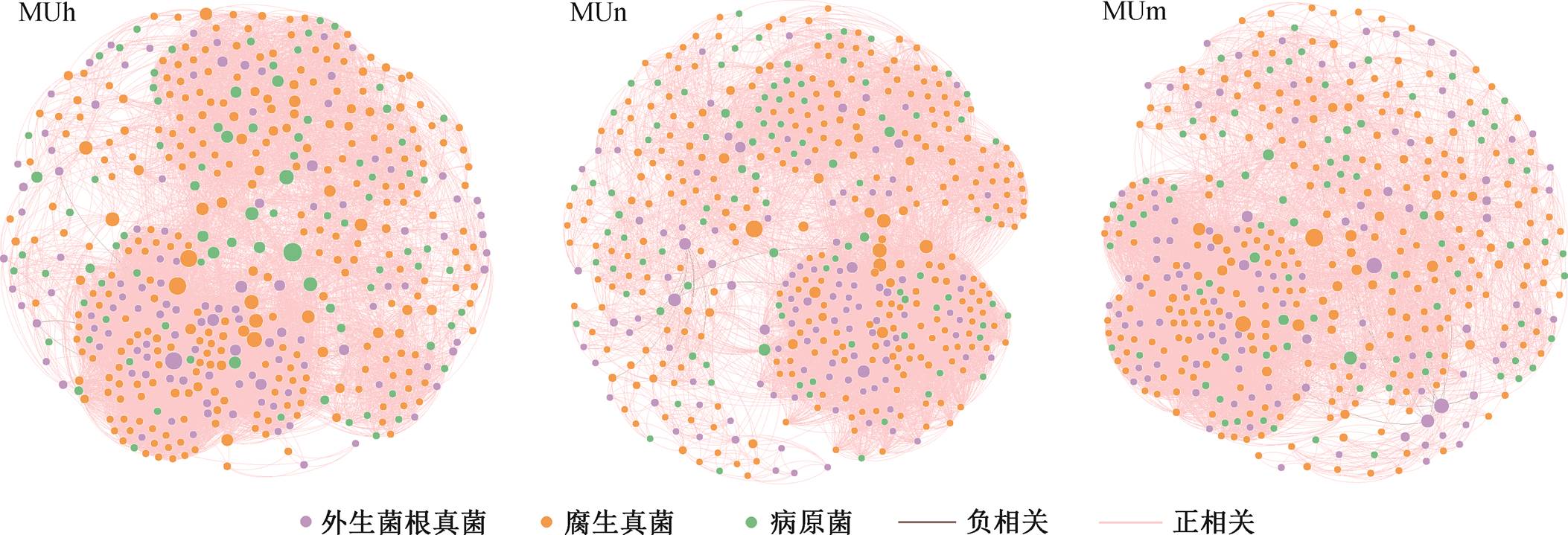
节点大小表示中介中心性大小, 下同
图1 毛乌素沙地不同林龄樟子松人工林根内真菌群落共现网络
Fig. 1 Co-occurrence network of fungal community in roots of P. sylvestris plantation of different stand ages in the Mu Us Desert
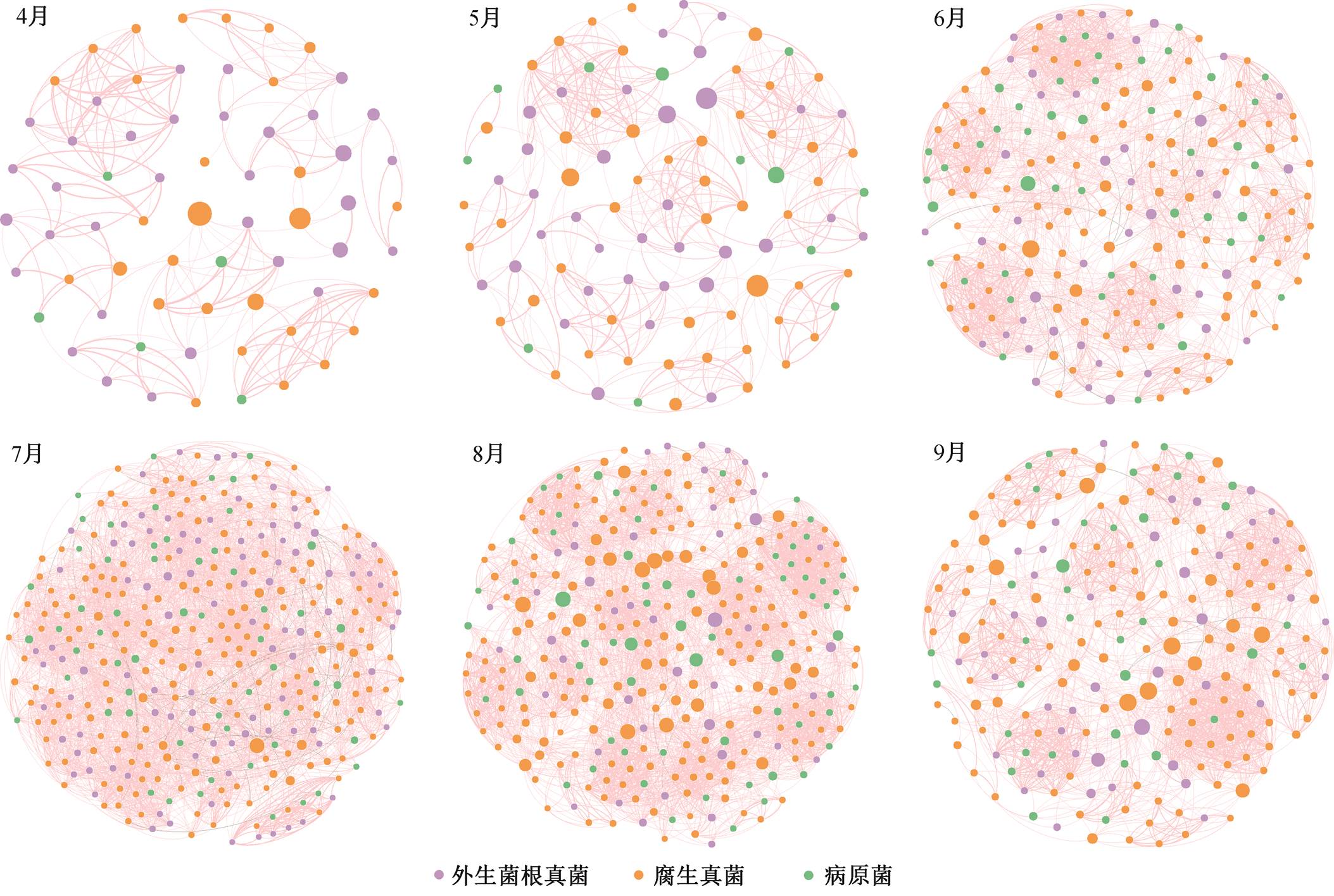
图2 毛乌素沙地不同月份樟子松人工林根内真菌群落共现网络
Fig. 2 Co-occurrence network of fungal community in root of P. sylvestris plantation of different months in the Mu Us Desert
选择根内真菌网络中介中心排前 10 位的 OTU, 并结合节点拓扑角色, 筛选在根内真菌网络中起重要作用的关键菌种(表 4)。其中, 生长季初期共有12 个关键属, 主要为外生菌根真菌, 包括地孔菌属(Geopora)和棉革菌属(Tomentella)等; 生长季旺盛期共有 8 个关键属, 主要为腐生真菌和病原真菌, 包括暗球腔菌属(Phaeosphaeria)、Neocucurbitaria、被孢霉属(Mortierella)、篮状菌属(Talaromyces)、红酵母属(Rhodotorula)以及新丽鞭菌属(Neocalli- mastix); 生长季末期共有 8 个关键属, 主要为腐生真菌, 包括地孔菌属和篮状菌属等。近熟林中未筛选出共有属; 中龄林共有 2 个关键属, 分别是 Neo-cucurbitaria 和瓶毛壳属(Lophotrichus); 成熟林共有3 个共有属, 分别是块菌属(Tuber)、篮状菌属和被孢霉属。
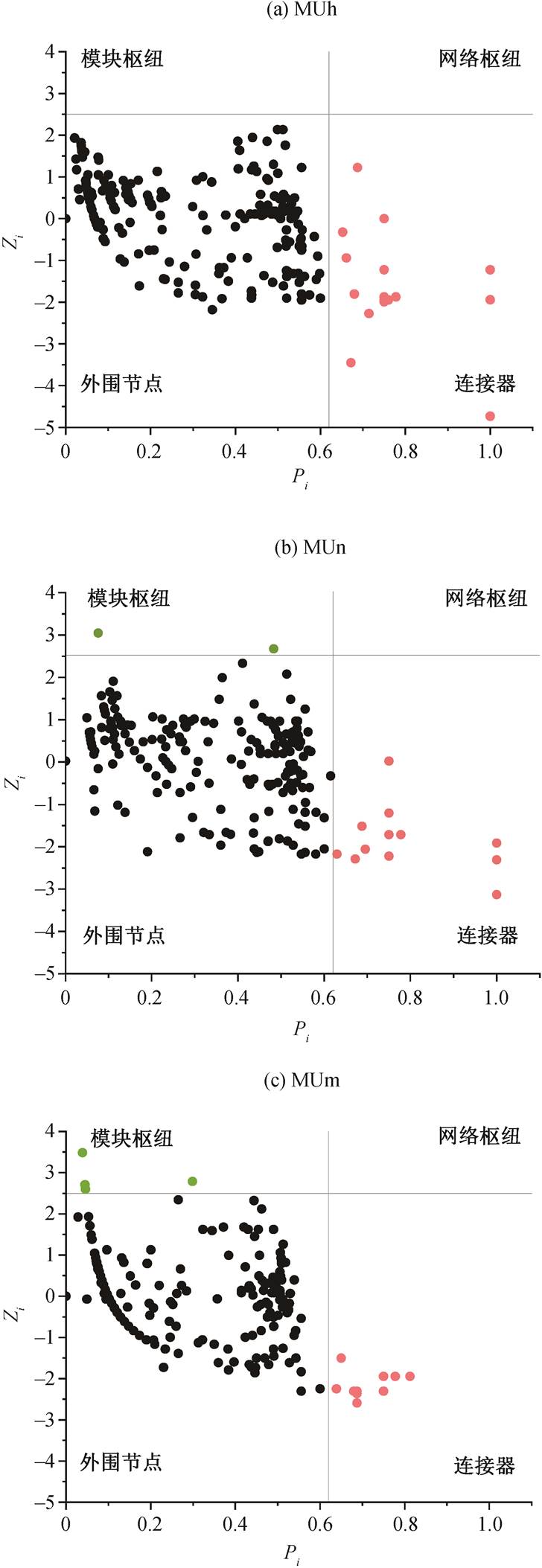
图3 毛乌素沙地不同林龄樟子松人工林根内真菌拓扑角色
Fig. 3 Fungal topological roles in roots of P. sylvestris plan-tation of different stand ages in the Mu Us Desert
表4 毛乌素沙地樟子松人工林根内真菌关键菌种
Table 4 Key fungal species in roots of P. sylvestris plantation in the Mu Us Desert
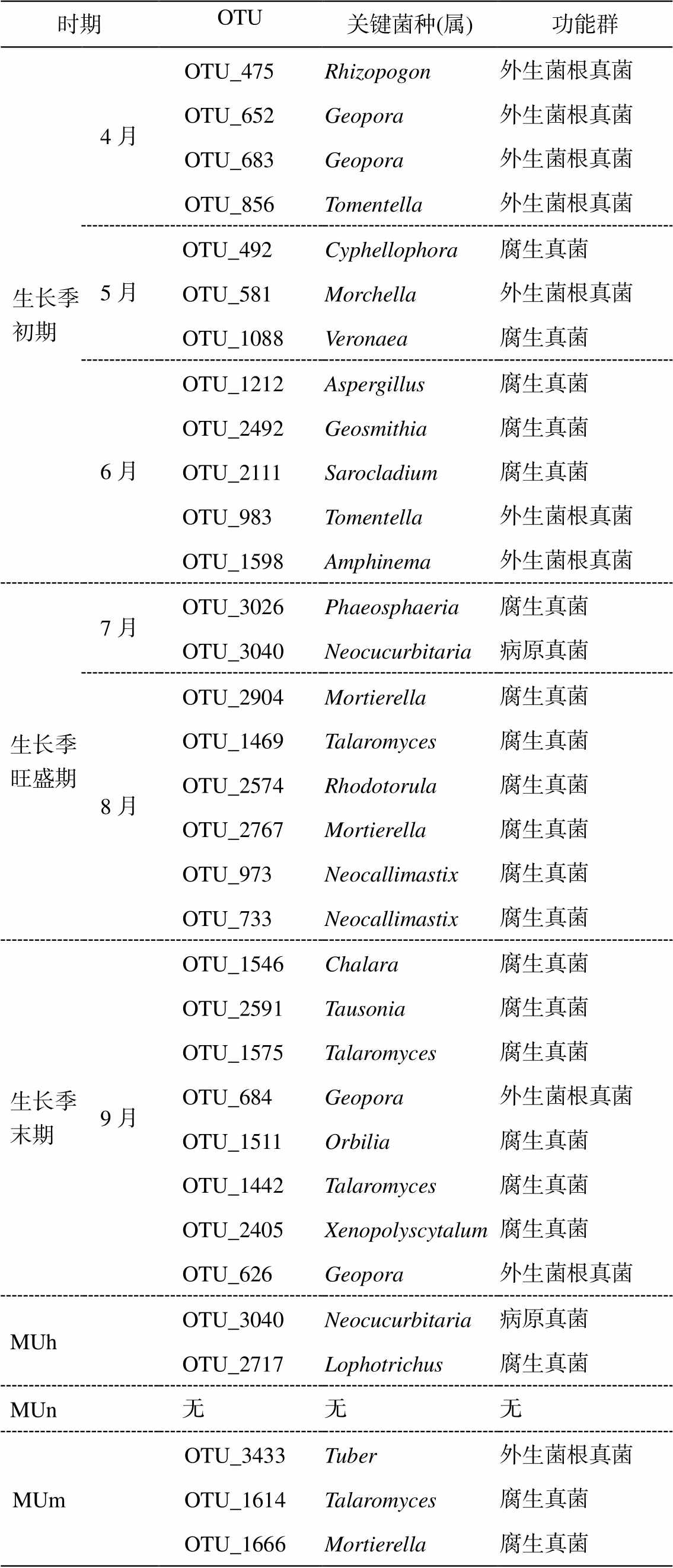
时期OTU关键菌种(属)功能群 生长季初期4月OTU_475Rhizopogon外生菌根真菌 OTU_652Geopora外生菌根真菌 OTU_683Geopora外生菌根真菌 OTU_856Tomentella外生菌根真菌 5月OTU_492Cyphellophora腐生真菌 OTU_581Morchella外生菌根真菌 OTU_1088Veronaea腐生真菌 6月OTU_1212Aspergillus腐生真菌 OTU_2492Geosmithia腐生真菌 OTU_2111Sarocladium腐生真菌 OTU_983Tomentella外生菌根真菌 OTU_1598Amphinema外生菌根真菌 生长季旺盛期7月OTU_3026Phaeosphaeria腐生真菌 OTU_3040Neocucurbitaria病原真菌 8月OTU_2904Mortierella腐生真菌 OTU_1469Talaromyces腐生真菌 OTU_2574Rhodotorula腐生真菌 OTU_2767Mortierella腐生真菌 OTU_973Neocallimastix腐生真菌 OTU_733Neocallimastix腐生真菌 生长季末期9月OTU_1546Chalara腐生真菌 OTU_2591Tausonia腐生真菌 OTU_1575Talaromyces腐生真菌 OTU_684Geopora外生菌根真菌 OTU_1511Orbilia腐生真菌 OTU_1442Talaromyces腐生真菌 OTU_2405Xenopolyscytalum腐生真菌 OTU_626Geopora外生菌根真菌 MUhOTU_3040Neocucurbitaria病原真菌 OTU_2717Lophotrichus腐生真菌 MUn无无无 MUmOTU_3433Tuber外生菌根真菌 OTU_1614Talaromyces腐生真菌 OTU_1666Mortierella腐生真菌
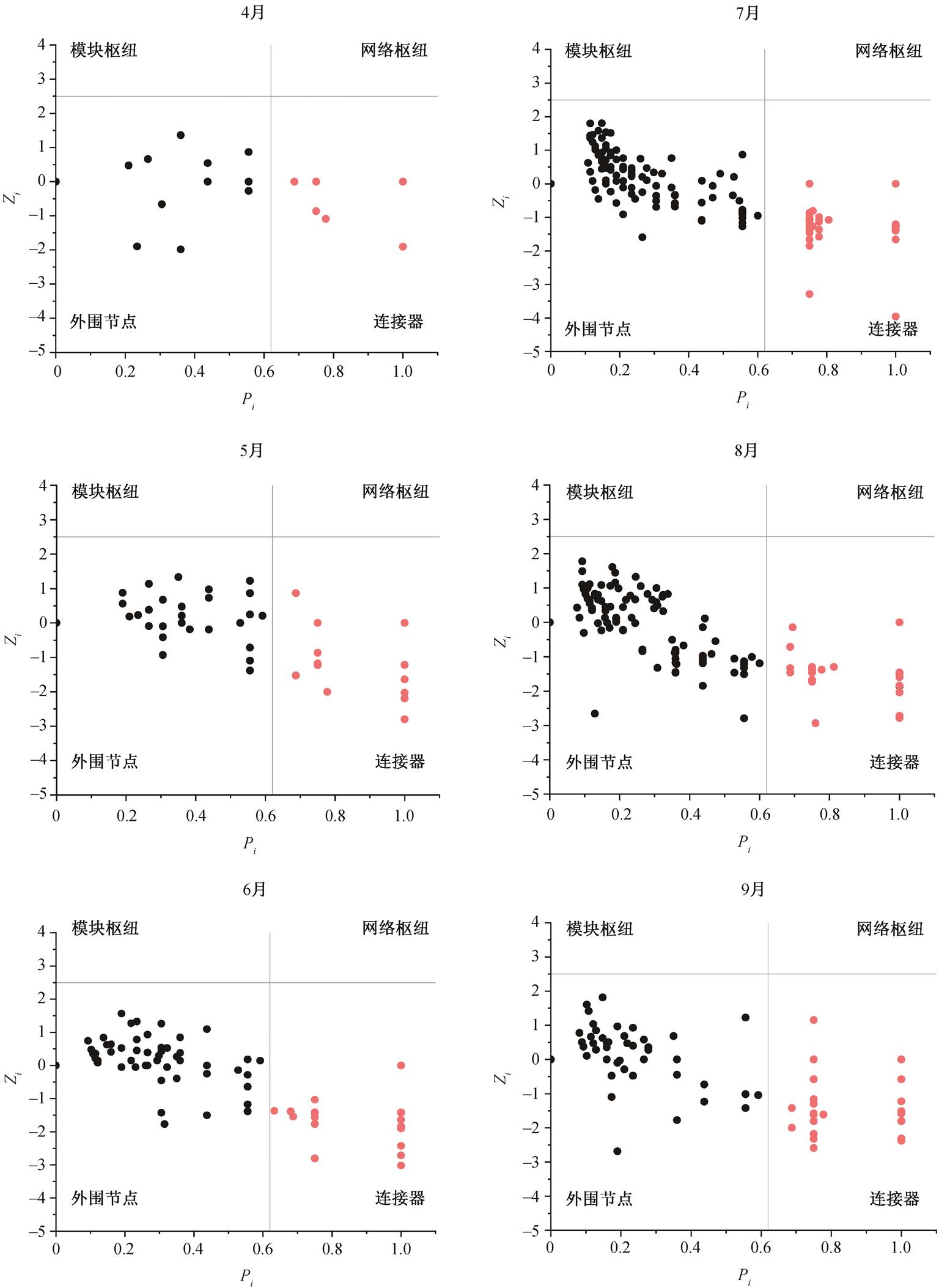
图4 毛乌素沙地不同月份樟子松人工林根内真菌拓扑角色
Fig. 4 Fungal topological roles in roots of P. sylvestris plantation of different months in the Mu Us Desert
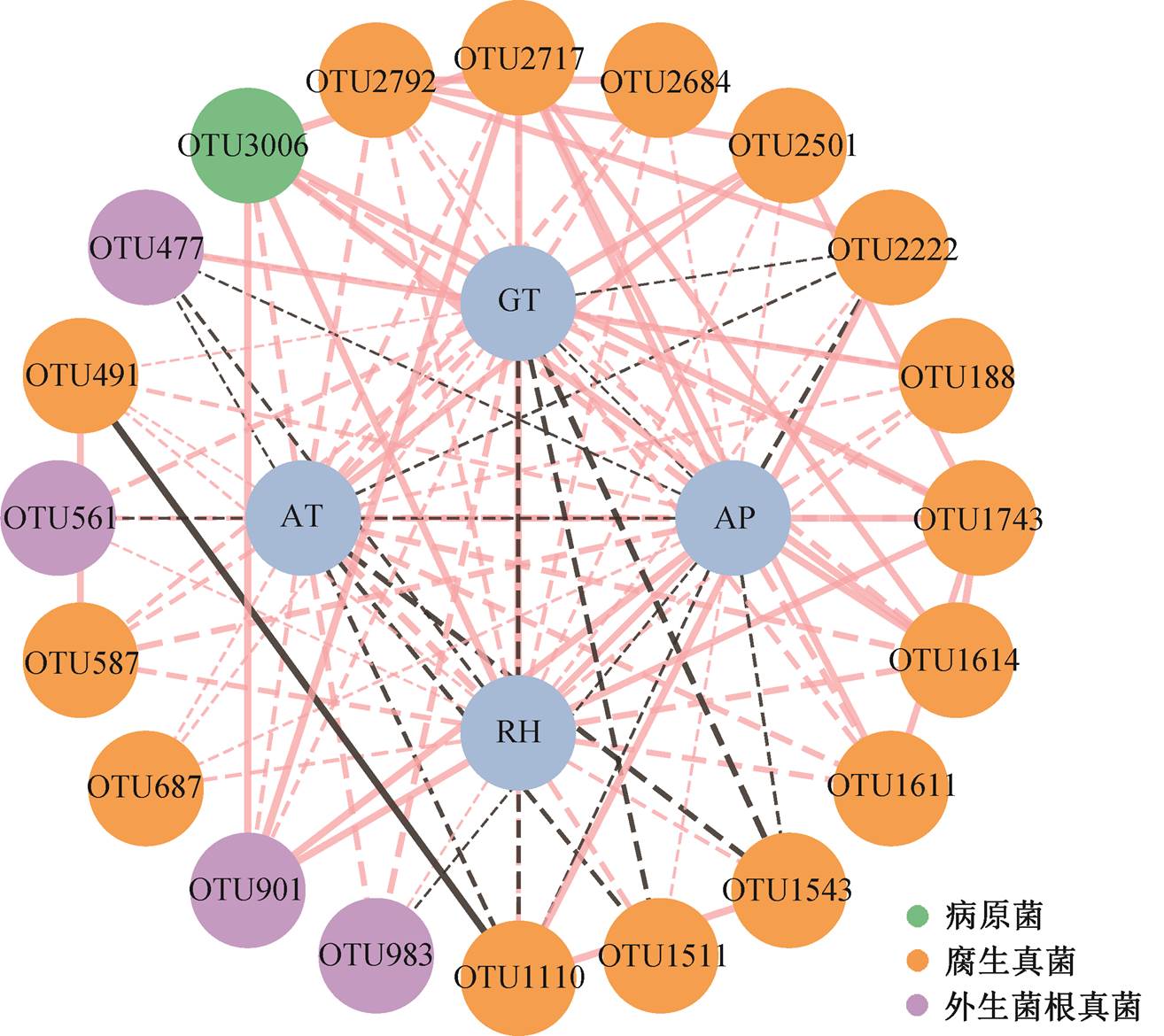
虚线表示相关性不显著。AT:气温; GT:地温; AP:降水; RH:相对湿度
图5 毛乌素沙地樟子松人工林根内真菌群落与气候因素网络关系
Fig. 5 Relationship between fungal community and climate factors in roots of P. sylvestris plantation in the Mu Us Desert
选择樟子松根内真菌网络中介中心性最高的 20 个 OTU, 与气象因子建立网络关系(图 5), 可以看出, 气温和地温均显著地影响腐生真菌 OTU2501, 平均降水量和相对湿度在网络中与 OTU 有最多的显著相关关系(P<0.05)。将不同功能群的根内真菌与环境因子 Mantel 进行检验(表 5), 可以发现, 总网络与相对湿度呈极显著相关(P<0.01)。平均降水量显著地影响腐生真菌网络和病原真菌网络, 相对湿度显著地影响外生菌根真菌网络和病原真菌网络(P<0.05), 并与腐生真菌网络极显著相关(P<0.01)。
毛乌素沙地樟子松人工林根内真菌群落网络随林龄的增加变化不明显, 可能是宿主根系分泌物招募策略和土壤生境的改变导致群落网络构建的动态变化[21]。不同生长季阶段则存在明显的差异, 气候的季节性变化更强烈地影响根内真菌网络。樟子松人工林根内真菌群落网络在生长季初期和末期的节点数和边数明显低于生长季旺盛期, 网络更加简单, 物种之间孤立, 可能是由于水分和养分的可利用性降低, 造成资源限制增强, 导致微生物多样性和网络复杂性降低[22]。腐生真菌和病原真菌的丰度在生长季旺盛期达到最大值, 远高于外生菌根真菌, 樟子松易受植物病害[23]。
表5 毛乌素沙地樟子松人工林根内真菌网络与气候因子中介中心性的Mantel检验结果
Table 5 Mantel test for betweenness centrality between fungal network and climatic factors in P. sylves-tris plantation in the Mu Us Desert

真菌网络气温地温降水相对湿度 总网络0.2460.3570.0210.729** 外生菌根真菌网络0.6920.5950.6760.743* 腐生真菌网络0.6830.5810.898*0.940** 病原真菌网络0.7990.7180.858*0.865*
说明: * P<0.05, ** P<0.01。
本研究中, 不同林龄和不同生长季阶段共现网络的正相关性均多于负相关性, 物种之间合作与生态位重叠, 与前人对樟子松林土壤微生物群落的研究结果[24]一致。樟子松人工林根内真菌群落以协同合作为主, 竞争关系较弱, 主要通过物种间的合作来抵抗干旱半干旱地区较为恶劣的环境条件[14,25]。此外, 生长季旺盛期负相关最多, 竞争加强, 主要原因是此阶段病原真菌的增多及其活性的增强, 外生菌根真菌和腐生真菌对病原真菌直接或间接的拮抗作用也相应地增强[18]。
平均降水量和相对湿度是影响毛乌素沙地樟子松林根内真菌群落网络的主要因素。真菌网络对降水的季节性变化敏感, 这与亚热带森林土壤真菌网络的研究结果[13]一致。7—8 月, 毛乌素沙地降水比其他月份多, 利于植物生长, 是樟子松的生长季旺盛期。降水增加了土壤含水量, 导致土壤相通过液压连接变得更紧密, 生态位分化较弱, 进而增强了生长季旺盛期的樟子松外生菌根真菌、腐生真菌和病原真菌之间的相互作用, 根内真菌网络比其他时期更加复杂[26]。樟子松生长季初期和末期, 毛乌素沙地降水量小, 根内真菌群落则需要通过更多的正相互作用来缓解环境的胁迫, 同时扩大自己的生态位, 以便更好地获取资源[27]。此外, 本研究中樟子松根内真菌群落网络与相对湿度显著相关(P<0.05)。相对湿度受温度和降水共同调控, 在较温暖的环境中, 微生物分类群之间的相互作用相应地增强[28]。毛乌素沙地 7—8 月相对湿度较高, 促进樟子松气孔张开, 且多数腐生真菌更偏好湿润环境, 根内真菌群落增强相互作用, 以便对植物养分的利用[29–30]。同时, 根内真菌生物量、多样性以及土壤养分随之增加, 为相互作用提供更多的机会。
在不同林龄和生长季阶段, 樟子松根内真菌关键菌种存在动态变化, 关键种群在维持网络稳定性中发挥重要作用, 与群落的功能密切联系[31]。随着生长季的推移, 主要营养型由外生菌根真菌变为腐生真菌。生长季初期, 与菌根共生的樟子松向根外释放的碳源大部分优先分配给菌根真菌, 外生菌根真菌促进樟子松吸收养分和水分, 成为主要关键菌种[32]。地孔菌属和棉革菌属是针叶林中常见且占主导地位的外生菌根真菌类群, 能高度适应盐碱干旱环境[33]。生长季末期, 凋落物、动植物残体的增加促使作为分解者的腐生真菌成为主要关键菌种。生长季旺盛期, 樟子松根内真菌网络更加复杂, 关键菌种在生长季旺盛期的作用便显得尤为重要。
生长季旺盛期, 樟子松根内真菌群落网络的关键菌种在网络中的节点数较少, 且相对丰度低, 表明稀有类群在樟子松根内真菌群落中也可能发挥关键作用。营养型为腐生真菌和病原真菌, 其中暗球腔菌属常腐生于单子叶植物的枯死茎或其他木材上, 此外, 暗球腔菌属也有潜在的病原性, 作为有效的生防菌, 对马唐(Digitaria sanguinalis)具有强致病性[34]; 篮状菌是适应性非常强的腐生真菌, 可帮助植物抑制病原真菌, 如黄色篮状菌(T. flavus)对尖孢镰刀菌(Fusarium oxysporum)等病原真菌有拮抗作用[35]; 红酵母属多被认为是一种拮抗酵母菌, 可与病原真菌竞争植物表面营养及侵染位点, 形成保护屏障, 或与病原真菌竞争空间条件, 达到抑制病害的目的[36]。被孢霉属是丝状腐生真菌, 可产生多种不饱和脂肪酸, 对某些病原真菌有抑制作用[37]。值得注意的是, Neocucurbitaria 大部分为致病菌或植物寄生菌, 易引发叶斑病等植物病害, 然而在樟子松根内真菌网络中, 它作为关键菌种, 与其他根内真菌并不存在竞争或拮抗关系, 可能是由于气候环境的改变, Neocucurbitaria 表现出不同的生存策略, 以腐生的营养方式生活[38–39]。此外, 磷是沙地樟子松林生长的重要限制因子, 植物根部只能吸收可溶性磷, 而被孢霉是一种解磷真菌, 更具侵染繁殖优势, 前人对施用磷肥后砂姜黑土真菌的研究结果也表明, 腐生真菌具有较高的竞争力, 可以满足不同生态位的真菌生长[19,40]。
在针叶树种凋落物分解过程中, 真菌群落需面对更多的根系分泌物以及含量较高且难分解的半乳甘露聚糖, 因此在不同林龄和生长季阶段, 腐生真菌在土壤中具有较高的竞争力, 对维持真菌网络稳定性均有重要作用[29]。樟子松在中龄林阶段即处于快速生长时期, 凋落物和根系分泌物的输入量随着林龄的增加而增加, 因此腐生真菌发挥着关键作用[19]。在生长季旺盛期, 有益的腐生真菌作为关键类群, 在复杂的真菌网络中起着关键作用, 通过诱导樟子松免疫、分泌抗生素或与病原真菌竞争资源来促进樟子松健康生长[41]。
1)毛乌素沙地不同林龄阶段樟子松人工林根内真菌群落共现网络不存在明显的差异; 随着生长季推移, 根内真菌共现网络存在明显的差异, 根内真菌群落的相互作用先增强后减弱。毛乌素沙地樟子松人工林根内真菌以协同合作为主, 生长季旺盛期网络比其他生长季阶段更加复杂。
2)樟子松中龄林的关键类群为腐生真菌和病原真菌, 成熟林为外生菌根真菌和腐生真菌。生长季初期和末期的关键类群为外生菌根真菌和腐生真菌, 生长季旺盛期的关键类群为腐生真菌和病原真菌。在不同林龄和不同生长季阶段, 关键菌种主要为地孔菌属、棉革菌属、暗球腔菌属、Neocucurbi-taria 和被孢霉属等。
3)樟子松人工林根内真菌群落网络的影响因子为相对湿度, 外生菌根真菌受相对湿度影响, 腐生真菌网络和病原真菌网络受平均降水量和相对湿度影响。较高降水量和湿度的生长季旺盛期, 关键类群为腐生真菌和病原真菌, 对樟子松的生长和健康状况有着潜在的影响。
参考文献
[1] Baldrian P. Forest microbiome: diversity, complexity and dynamics. FEMS Microbiology Reviews, 2017, 41(2): 109–130
[2] Genre A, Lanfranco L, Perotto S, et al. Unique and common traits in mycorrhizal symbioses. Nature Re-views Microbiology, 2020, 18: 649–660
[3] Anthony M A, Frey S D, Stinson K A. Fungal commu-nity homogenization, shift in dominant trophic guild, and appearance of novel taxa with biotic invasion. Ecosphere, 2017, 8(9): e01951
[4] 马泊泊, 黄瑞林, 张娜, 等. 秸秆生物质炭对根际土壤细菌–真菌群落分子生态网络的影响. 土壤学报, 2019, 56(4): 964–974
[5] Fuhrman J A. Microbial community structure and its functional implications. Nature, 2009, 459: 193–199
[6] Nilsson R H, Anslan S, Bahram M, et al. Mycobiome diversity: high-throughput sequencing and identifica-tion of fungi. Nature Reviews Microbiology, 2019, 17: 95–109
[7] Barberan A, Bates S T, Casamayor E O, et al. Using network analysis to explore co-occurrence patterns in soil microbial communities. The ISME Journal, 2012, 6: 343–351
[8] Karimi B, Dequiedt S, Terrat S, et al. Biogeography of soil bacterial networks along a gradient of cropping intensity. Scientific Reports, 2019, 9: 3812
[9] Zhou J, Deng Y, Luo F, et al. Functional Molecular Ecological Networks. mBio, 2010, 1(4): e00169 -10
[10] Zhao P S, Gao G L, Ren Y, et al. Intra-annual varia-tion of root-associated fungi of Pinus sylvestris var. mongolica: the role of climate and implications for host phenology. Applied Soil Ecology, 2022, 176: 104480
[11] Zhang Y H, Ni J, Tang F P, et al. Root-associated fungi of Vaccinium carlesii in subtropical forests of China: intra- and inter-annual variability and impacts of human disturbances. Scientific Reports, 2016, 6: 22399
[12] Yuan M M, Guo X, Wu L, et al. Climate warming enhances microbial network complexity and stability. Nature Climate Change, 2021, 11: 343–348
[13] He D, Shen W, Eberwein J, et al. Diversity and co-occurrence network of soil fungi are more responsive than those of bacteria to shifts in precipitation sea-sonality in a subtropical forest. Soil Biology and Bio-chemistry, 2017, 115: 499–510
[14] Zhou H, Gao Y, Jia X H, et al. Network analysis reveals the strengthening of microbial interaction in biological soil crust development in the Mu Us Sandy Land, northwestern China. Soil Biology and Bioche-mistry, 2020, 144: 107782
[15] Sokol N W, Slessarev E, Marschmann G L, et al. Life and death in the soil microbiome: how ecological processes influence biogeochemistry. Nature Reviews Microbiology, 2022, 20: 415–430
[16] Tao J M, Meng D, Qin C, et al. Integrated network analysis reveals the importance of microbial interac-tions for maize growth. Applied Microbiology and Biotechnology, 2018, 102(8): 3805–3818
[17] 丁钰珮, 杜宇佳, 高广磊, 等. 呼伦贝尔沙地樟子松人工林土壤细菌群落结构与功能预测. 生态学报, 2021, 41(10): 4131–4139
[18] 赵珮杉, 郭米山, 高广磊, 等. 科尔沁沙地樟子松根内真菌群落结构和功能群特征. 林业科学, 2020, 56(9): 87–96
[19] 柳叶, 任悦, 高广磊, 等. 沙地樟子松人工林土壤碳氮磷储量分布特征. 中国水土保持科学, 2021, 19(6): 27–34
[20] 魏晓帅, 郭米山, 高广磊, 等. 呼伦贝尔沙地樟子松根内真菌群落结构与功能群特征. 北京大学学报(自然科学版), 2020, 56(4): 710–720
[21] Lucasborja M E, Hedo J, Cerda A, et al. Unravelling the importance of forest age stand and forest structure driving microbiological soil properties, enzymatic activities and soil nutrients content in Mediterranean Spanish black pine (Pinus nigra Ar. ssp. salzmannii) Forest. Science of the Total Environment, 2016, 562: 145–154
[22] Banerjee S, Walder F, Büchi L, et al. Agricultural intensification reduces microbial network complexity and the abundance of keystone taxa in roots. The ISME Journal, 2019, 13: 1722–1736
[23] Chu H, Wang C, Wang H, et al. Pine wilt disease alters soil properties and root-associated fungal com-munities in Pinus tabulaeformis forest. Plant and Soil, 2016, 404(1/2): 237–249
[24] Bi B Y, Zhang H, Yuan Y, et al. Dynamic changes of soil microbial community in Pinus sylvestris var. mongolica plantations in the Mu Us Sandy Land. Journal of Environmental Management, 2021, 287: 112306
[25] Hoek T A, Axelrod K, Biancalani T, et al. Resource availability modulates the cooperative and competitive nature of a microbial cross-feeding mutualism. PLoS Biology, 2016, 14(8): 1002540
[26] Wang G, Or D. Hydration dynamics promote bacterial coexistence on rough surfaces. The ISME Journal, 2013, 7(2): 395–404
[27] Freilich M A, Wieters E, Broitman B R, et al. Species co-occurrence networks: can they reveal trophic and non-trophic interactions in ecological communities?. Ecology, 2018, 99(3): 690–699
[28] 刘亚玲, 信忠保, 李宗善, 等. 河北坝上樟子松人工林径向生长及其对气候因素的响应. 生态学报, 2022, 42(5): 1830–1840
[29] 李德会, 韩周林, 吴庆贵, 等. 影响森林细根分 解的腐生真菌功能特性研究综述. 世界林业研究, 2021, 34(3): 25–31
[30] 刘艳娇, 樊丹丹, 李香真, 等. 人工与天然云杉林土壤真菌群落多样性及菌群网络关系特征. 应用生态学报, 2021, 32(4): 1441–1451
[31] Li B B, Roley S S, Duncan D S, et al. Long-term excess nitrogen fertilizer increases sensitivity of soil microbial community to seasonal change revealed by ecological network and metagenome analyses. Soil Biology and Biochemistry, 2021, 160: 108349
[32] Kraft B, Strous M, Tegetmeyer H E. Microbial nitrate respiration-genes, enzymes and environmental distri-bution. Journal of Biotechnology, 2011, 155(1): 104–117
[33] 王家源, 殷小琳, 任悦, 等. 毛乌素沙地樟子松外生菌根真菌多样性特征. 微生物学通报, 2020, 47 (11): 3856–3867
[34] 邹德勇, 杨智越, 杜春梅. Phaeosphaeria sp. HD-06菌株对马唐的致病性分析及初步鉴定. 植物保护, 2020, 46(4): 98–104
[35] 孙剑秋, 阮永明, 金世宇, 等. 篮状菌属的重要性及其分类学研究概况. 菌物研究, 2021, 19(2): 83–93
[36] 于帅, 刘天明, 魏渱. 拮抗酵母菌对果蔬采后病害生物防治的研究进展. 食品工业科技, 2010(9): 402–405
[37] 谢尔瓦尼木·阿不都艾尼, 海孜尼姑力·努尔, 赵歉歉, 等. 枯草芽孢杆菌 J-15 抗大丽轮枝菌次生代谢产物对棉田土壤真菌多样性的影响. 微生物学通报, 2021, 48(6): 1997–2007
[38] Wanasinghe D N, Phookamsak R, Jeewon R, et al. A family level rDNA based phylogeny of Cucurbitaria-ceae and Fenestellaceae with descriptions of new Fenestella species and Neocucurbitaria gen. nov. Mycosphere, 2017, 8(4): 397–414
[39] Márquez S S, Bills G F, Herrero N, et al. Non-systemic fungal endophytes of grasses. Fungal Eco-logy, 2012, 5(3): 289–297
[40] 马垒, 赵文慧, 郭志彬, 等. 长期不同磷肥施用量对砂姜黑土真菌多样性、群落组成和种间关系的影响. 生态学报, 2019, 39(11): 4158–4167
[41] Gao M, Xiong C, Gao C, et al. Disease-induced changes in plant microbiome assembly and functional adaptation. Microbiome, 2021, 9: 187
Network Features of Root-Associated Fungi of Pinus Sylvestris var.Mongolica Plantations and Response to Climate Factors in the Mu Us Desert
Abstract To reveal the dynamic characteristics of root-associated fungal communities of P. sylvestris plantation, the co-occurrence network of root-associated fungi in P. sylvestris plantation were analyzed by using molecular ecological network analysis in the Mu Us Desert and its response to climate factors in different stand ages (27 a, 33 a and 44 a) and their annual growth stages (early growth season, vigorous growth season and end-of-growth season).The following conclusions are obtained. 1) Different growing seasons rather than different stand ages had significant differences in the co-occurrence network of fungi in roots of P. sylvestris plantation, and the co-occurrence network of fungi in roots was more complex in the vigorous growing season. 2) The keystone species of root-associated fungi of P. sylvestris plantation were significant in different growing seasons and stand ages. The keystone genera mainly included Geopora, Tomentella, Phaeosphaeria, Neocucurbitaria, Mortierella, etc. Ectomycorrhizal and saprophytic fungi were the keystone taxa in the early and end of the growth season, and saprophytic fungi and pathogens were the keystone taxa in the vigorous growth season. Meanwhile, the keystone taxa of middle-aged forest were saprophytic fungi and pathogens, while the mature forest were ectomycorrhizal and saprophytic fungi. 3) Relative humidity was the main influencing factor for ectomycorrhizal fungi network, and average precipitation and relative humidity were the main influencing factors for saprophytic fungi network and pathogens network (P<0.05).Therefore, the network dynamic of root-associated fungi of P. sylvestris plantation in different growing seasons was stronger than that of stand age in Mu Us Desert, and the co-occurrence network structure was more complex in the vigorous growth season. The root-associated fungal network was mainly affected by average precipitation and relative humidity. Saprophytic fungi played an important role in maintaining the stability of the fungal network in different stand ages and growing seasons. Saprophytic fungi and pathogens in the vigorous growth season with high precipitation and relative humidity, which potentially contributed to the growth and health of P. sylvestris plantation.
Key words psammophyte; stand age; growing season; soil fungi; co-occurrence network; climate factor