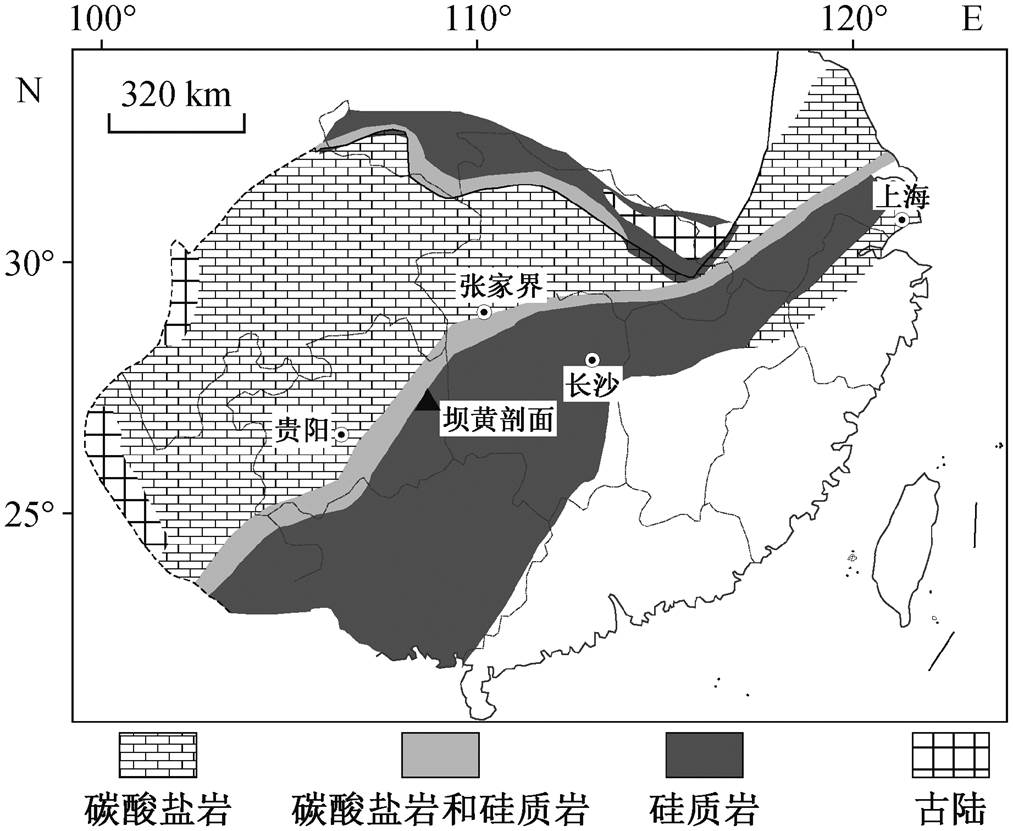
图1 埃迪卡拉纪–寒武纪扬子板块古地理图(修改自文献[17])
Fig. 1 Palaeogeographic map of the Yangtze Plate during the Ediacaran-Cambrian transition (modified from Ref. [17])
北京大学学报(自然科学版) 第59卷 第3期 2023年5月
Acta Scientiarum Naturalium Universitatis Pekinensis, Vol. 59, No. 3 (May 2023)
doi: 10.13209/j.0479-8023.2023.016
国家自然科学基金(41402025)、中国科学院南京地质古生物研究所现代古生物学和地层学国家重点实验室基金(133103)资助
收稿日期: 2022–05–02;
修回日期: 2022–05–17
摘要 对贵州省东部坝黄剖面留茶坡组硅质岩中的微体化石进行研究。通过薄片观察, 发现多种球形化石, 包括一类具连接体的球形化石以及两种具深色外壁的球形化石。对具连接体的球形化石进行系统的描述, 并建立新属 Bahuangia gen. nov.。Bahuangia的连接体呈葫芦状或链状, 形态可与出芽生殖的单细胞真菌或细菌进行类比。所报道的3类球形化石在扬子板块埃迪卡拉纪晚期的深水相地层中广泛分布, 此次发现加深了对埃迪卡拉纪晚期深水区生物面貌的认识, 可为区域地层对比以及地球早期生命演化的研究提供新的材料。
关键词 球形化石; 硅质岩; 留茶坡组; 贵州
埃迪卡拉纪–寒武纪时期经历了早期地球环境与生命的重大变革。自新元古代全球性冰期事件结束后, 这一时期记录了一系列环境变化事件, 包括海洋大规模氧化以及碳同位素的最大负漂移等[1–3]。伴随全球环境的改变, 地球生物圈也迎来重大更迭, 地球生命出现爆发式转变。该时期不仅是后生动物演化的起点, 也完成从微体生物为主的元古宙生物圈向以后生动物为代表的显生宙生物圈的转变, 奠定了显生宙生物圈发展的基础[3–4]。
埃迪卡拉纪晚期至寒武纪早期, 我国华南扬子板块广泛发育连续的海相地层, 其西北部浅水区以台地相碳酸盐岩为特征, 东南部深水区则表现为层状硅质岩[5–6]。前人已在扬子板块埃迪卡拉纪–寒武纪的地层内发现多个生物群, 是研究该时期生命演化的宝贵材料。然而, 这些生物群主要分布于浅水相区, 相对而言, 对深水相区生物特征的认识尚不充分。此外, 扬子板块埃迪卡拉系–寒武系界线的厘定也一直是研究重点。目前, 埃迪卡拉系–寒武系界线的全球界线层型位于加拿大纽芬兰的 Chapel Island 组, 并以遗迹化石 Trichophycus pedum 的首次出现为基准[7–8]。然而, 我国南方的浅水台地相通常以小壳化石的出现作为二者界线划分的标志。Yao 等[9]基于对塔里木盆地下寒武统微体化石组合的综合研究, 建立用于识别下寒武统底界的 Asteridium-Heliosphaeridium-Comasphaeridium (AHC) 疑源类化石带, 以小型具刺疑源类及管状化石 Megathrix 为特征, 可与扬子板块的微体化石组合对比[10–11]。然而, 鉴于化石材料的相对缺乏, 这一界线的识别在扬子板块深水相区依然遇到阻碍。
迄今为止, 在前寒武纪的硅质沉积中已发现多种生物化石[12–13], 硅化作用可使生物遗体以三维形态保存下来, 为研究地球早期生命演化提供重要窗口。本文选择贵州省东部坝黄村埃迪卡拉纪–寒武纪转折时期的留茶坡组硅质岩为研究对象, 对其中生物化石进行研究。在坝黄剖面留茶坡组中下部, 大量产出多种球形化石, 以孤立或连接体的形式出现。本文对一类具连接体的球形化石进行系统的描述, 并建立新属 Bahuangia gen. nov., 其形态可与以出芽方式进行繁殖的细菌或真菌类比。本文报道的三类球形化石在扬子板块深水区埃迪卡拉纪末期的地层中广泛分布[14–16], 可能代表该时期深水环境中发育的特征微体生物群落。本次研究丰富了扬子板块埃迪卡拉纪–寒武纪的生物化石种类, 可为深水区微体生物群落的组成提供新的材料, 同时对该时期区域地层的对比具有重要意义。
埃迪卡拉纪–寒武纪过渡时期, 扬子板块自西北向东南方向水体逐渐加深, 并依次形成台地、斜坡及盆地相[17–18](图 1)。该时期, 湖北、四川、重庆、陕南、云南和黔北等地区位于浅水区, 主要沉积以灯影组为代表的台地相碳酸盐岩; 湖南、桂北、黔东、黔南、皖南和浙西等深水区则以留茶坡组(对应安徽和江西的皮园村组以及广西的老堡组)互层的硅质岩和页岩为特征[5–6]。本文的研究材料为贵州省东部铜仁市坝黄剖面留茶坡组硅质岩(图2)。坝黄剖面留茶坡组厚约 6m, 主要表现为层状硅质岩和碳质页岩互层, 其上覆和下伏地层分别为牛蹄塘组页岩和陡山沱组页岩。在留茶坡组中部有一处火山凝灰岩层, 其 SIMS 锆石 U-Pb 年龄为542.6±3.7Ma[19], 对应埃迪卡拉纪晚期。
图1 埃迪卡拉纪–寒武纪扬子板块古地理图(修改自文献[17])
Fig. 1 Palaeogeographic map of the Yangtze Plate during the Ediacaran-Cambrian transition (modified from Ref. [17])
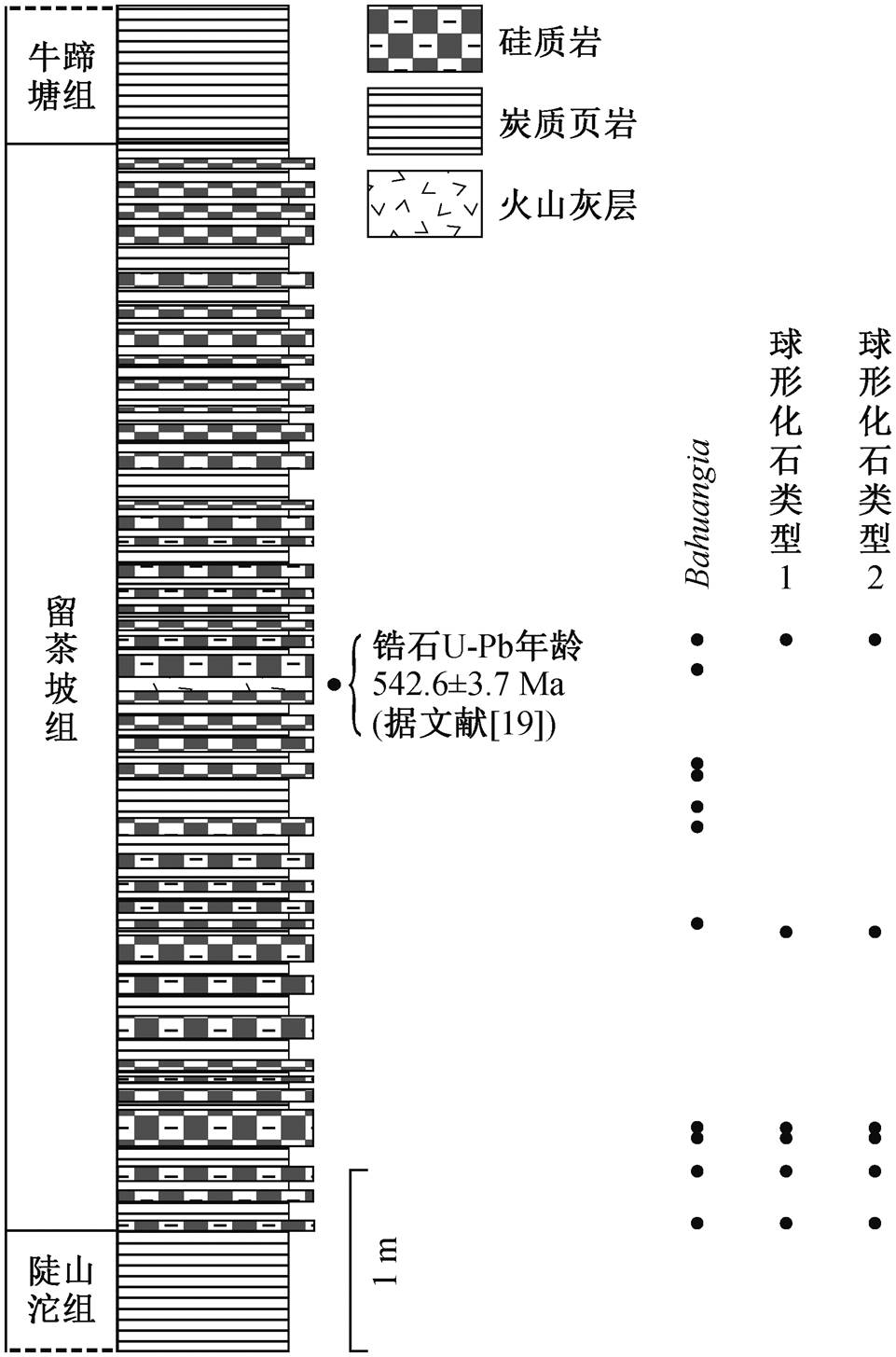
图2 贵州坝黄剖面留茶坡组柱状图
Fig. 2 Stratigraphic column of the Liuchapo Formation at Bahuang section in Guizhou Province
本文样品采自贵州省铜仁市坝黄村。主要利用岩石薄片, 对留茶坡组硅质岩中微体化石进行观察和描述, 化石观察均基于平行于层面的切片。薄片制作在北京大学薄片制备室完成, 在 Nikon LV100N POL 偏光显微镜下观察薄片, 使用 Nikon DS-Fi3 软件拍照, 并用 tpsDig 软件对化石进行测量。所有样品均保存于北京大学地质博物馆。
我们在贵州坝黄剖面留茶坡组硅质岩中发现一种中心呈颗粒状的球形化石(图 3(a)~(g)), 薄片中均呈圆形, 调整聚焦平面时, 直径有明显的变化, 因此判断其三维形态应为球形。该类球形化石分布于距剖面底部 0~3.2m 的区间内, 主要在剖面下部密集产出, 少量见于火山灰层之上。
这种球形化石具双层结构, 包括内质及外部晕圈(图 3)。外部晕圈较厚, 其外边缘平滑或有不规则突起。内质为具颗粒状的球形, 少数情况下不可辨认。球形化石的整体直径为 9~44μm (平均 21μm, n=32), 内质直径为 7~27μm (平均 16μm, n=32), 二者呈较好的正相关关系(图 4)。该类球形化石以孤立或群体的形式保存, 且球体之间常彼此连接(图 3 (c)~(g)), 表现为两个或多个球体以宽度不等的颈部相接, 形成葫芦状或链状(图 3(c)~(f))。此外, 还观察到一个具较长丝状连接物的标本(图 3(g)中黄色箭头所示)。鉴于其特殊的连接体形态与已知化石之间的区别, 本文为该球形化石建立新属 Bahuan-gia gen. nov. (系统描述见第 5 节)。
此外, 在坝黄剖面的中下部, 还发现一类具深色内质的球形化石(简称类型 1, 图 5 (a)~(c)), 以及一类仅保留外壁的球形化石(简称类型 2, 图 5(d))。
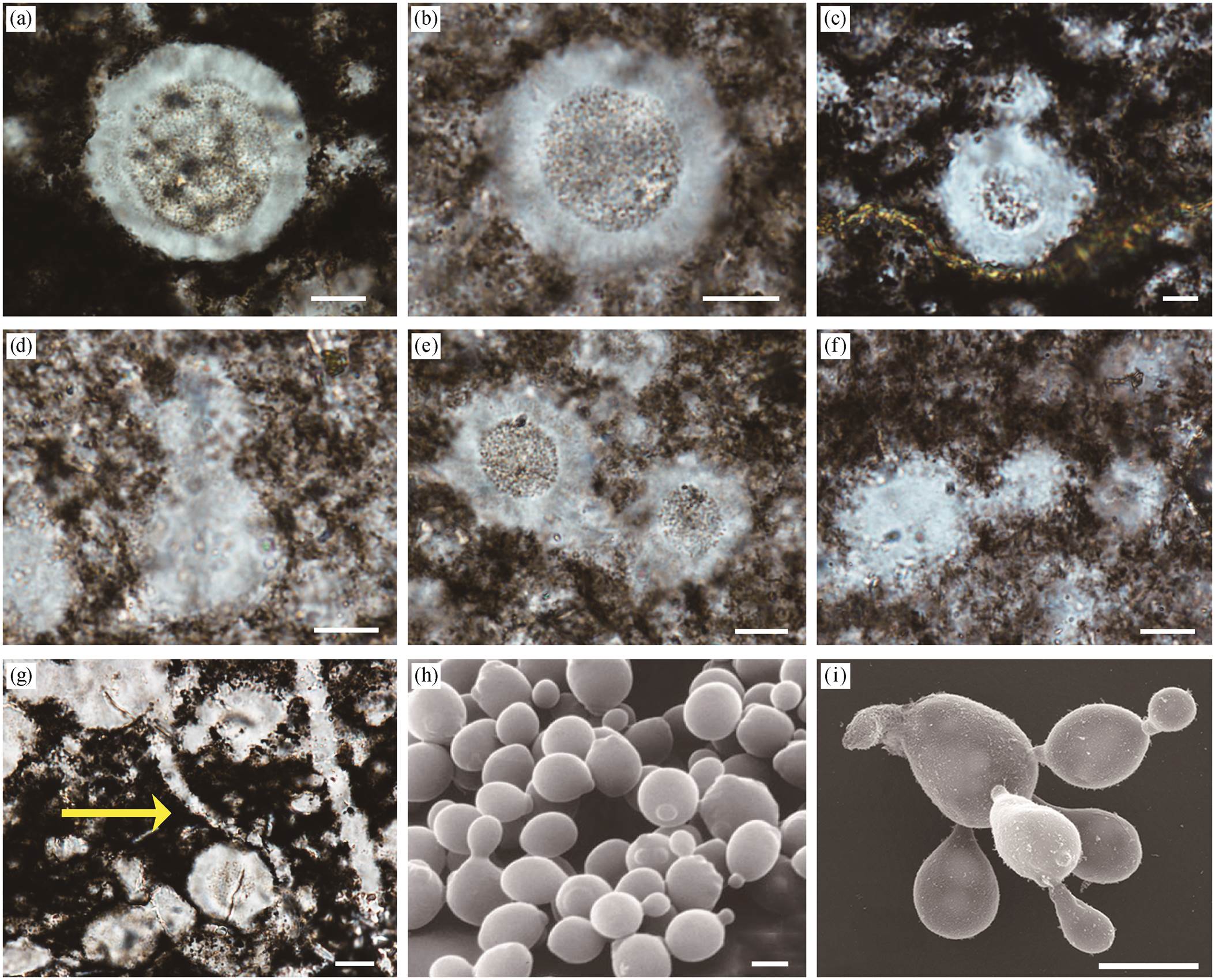
(h)和(i)据文献[18–19]。比例尺: (a)~(f)中代表10 μm; (g)中代表20 μm; (h)和(i)中代表5 μm
图3 坝黄剖面留茶坡组中的球形化石 Bahuangia gen. nov. ((a)~(g))以及出芽酵母菌((h)和(i))
Fig. 3 Bahuangia gen. nov. in the Liuchapo Formation at Bahuang section ((a)–(g)) and budding yeast ((h) and (i))
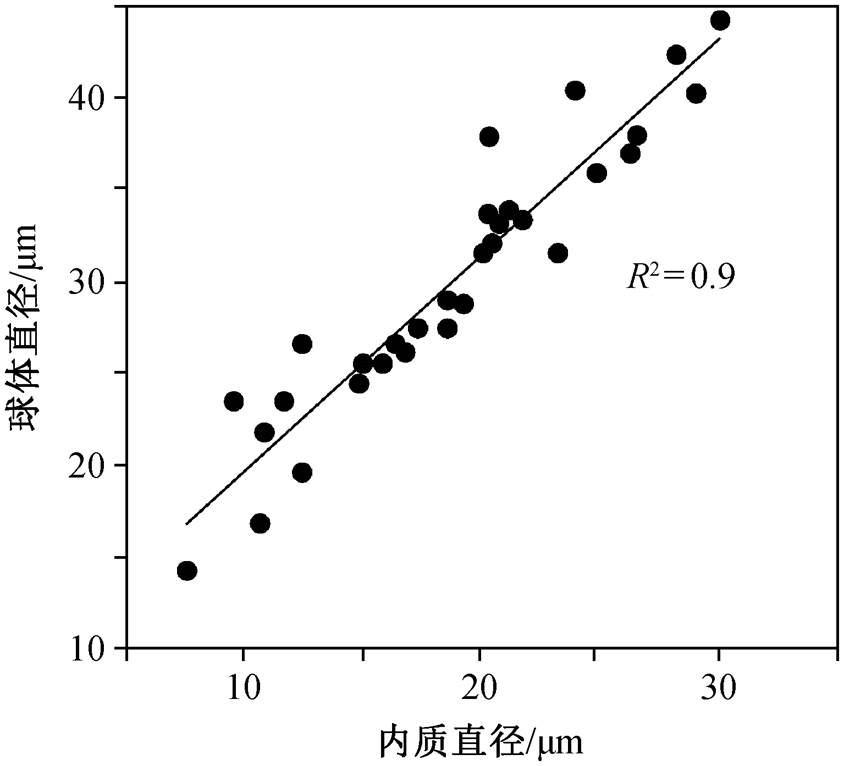
图4 具连接体球形化石 Bahuangia gen. nov.内质与球体直径
Fig. 4 Measurements of the spherical fossil Bahuangia gen. nov.
类型 1 具球形内质以及一较厚的深色外圈层, 二者成分相同, 以浅色硅质环带相隔。其深色外圈层可能是生物的原生结构, 或有机质降解过程中扩散侵染的结果。类型 1 的标本中, 外圈层厚度不稳定, 且边缘不清晰, 似乎更支持后一种解释; 介于内质与外圈层间的浅色环带, 边界较清晰, 可能代表生物的原始外壁。此外, 在类型 1 的球体外表面可观察到不均匀分布的突起。这种突起在每个标本中稳定存在, 应该为生物体本身的特征。基于标本保存状态的优劣, 有些突起呈规则的锥形(图 5(b)), 有些则较难辨认(图 5(c))。这种形态差异可能与埋藏过程中有机质的降解程度有关。类型 1 外圈层的整体直径为 16~40μm (平均 30μm, n=26), 内质直径为 9~26μm (平均 19μm, n=26), 外壁(浅色环带)厚0.9~3.7μm (平均 1.9μm, n=26)。根据其突起特征, 类型 1 可能为某种具刺疑源类。
类型 2 仅保存深色外壁, 其外壁轮廓不光滑, 内部为浅色的硅质填充。类型 2 直径为 37~76μm (平均 57μm, n=15), 外壁厚 6~9μm (平均 7.7μm, n=15)。
坝黄剖面中发现的球形化石 Bahuangia gen. nov., 其连接体形态与出芽生殖非常相似(图 6)。出芽生殖是一种无性生殖方式, 常见于真菌或细菌中, 也见于特定低等动物(如刺胞动物)中[20]。在真菌或细菌的出芽过程中, 首先由母体细胞一侧向外凸出, 形成芽基, 芽基与母细胞形态相似, 仅尺寸较小。芽基成熟之前, 保持与母细胞的连接, 并逐渐发育成完全复制母细胞的有机体。发育成熟后, 芽基脱离母细胞, 形成新个体。有时, 新形成的子细胞可能在与母细胞分离之前就开始新的出芽过程, 即在芽基上长出新的芽基。通过重复上述过程, 则可形成一系列细胞串。
考虑到连接体与出芽生殖方式的相似性, Bahu-angia gen. nov.可与某种单细胞细菌或真菌类比。例如, 同一连接体中, 球体尺寸总是存在明显的差异, 其中较大的球体可能为母细胞, 较小的则为发育中的芽基。球形酵母菌出芽时, 母体上可能连接着多个未脱落的芽基或子细胞, 类似葫芦状和链状等[21–22](如图 3(h)和(i)所示), 与本文观察到的连接方式极为类似。在出芽生殖初期, 母细胞体壁外会先形成小突起, 这也可以解释球形化石外圈层上不均匀的突起结构。因此, 坝黄剖面球形化石的多种保存形态可以恢复为出芽生殖过程的不同阶段(图6), 包括外壁小突起(开始出芽)、葫芦状或链状连接体(芽基生长)等。
丝状连接物仅见于一枚标本中(图 3(g)中黄色箭头所示), 可能与出芽生殖过程中产生的假菌丝有关。一些单细胞真菌在特定情况(如营养受限)下, 可发生细胞形态和分裂模式的变化[23], 其中包括一种假菌丝生长模式。以球形酵母菌为例, 在该生长模式下, 球形细胞拉长变细, 多个细胞之间通过细胞壁中的蛋白质保持连接, 形成假菌丝[24]。此外, 在加拿大北部中元古代 Grassy Bay 组中, 曾报道一种真菌化石 Ourasphaira giraldae[25], 其中也有两个球体以细丝相连(文献[25]中 Fig.1c), 其形态和大小均与坝黄剖面的标本类似。在 O. giraldae中, 末端的球体被类比为孢子囊, 而丝状体则被解释为菌丝。尽管菌丝与假菌丝均为细长的丝状结构, 二者之间依然存在明显的形态差异。例如, 假菌丝由一系列细胞串组成, 细胞之间无隔膜, 连接处有明显的缢缩; 菌丝细胞之间则有隔膜, 连接处没有宽度变化。然而, 由于坝黄剖面中仅发现一枚具丝状结构的标本, 且在丝状体中未保存更多的细节特征, 故不能做进一步的对比。
综合上述观察, 坝黄剖面留茶坡组中下部产出多种球形化石, 包括具连接体球形化石 Bahuangia gen. nov.以及两类具外壁的未定名球形化石。其中, Bahuangia gen. nov.可能为某种发育于深水的球形细菌或真菌, 其外壁的不规则突起以及连接体与出芽等无性生殖过程类似。结合坝黄剖面中部火山凝灰岩层的锆石 U-Pb 年龄(542.6±3.7Ma[19]), 本文描述的球形化石主要分布于埃迪卡拉纪晚期地层。此外, 与 Bahuangia gen. nov.类似的化石在贵州[14]和湖南[15]的留茶坡组下部均有发现, 本文描述的其他两类未定名球形化石也曾发现于贵州[14]和湖南[15]的留茶坡组以及安徽皮园村组[16]中。这些球形化石集中分布于埃迪卡拉纪晚期地层, 且在华南地区的深水相带广泛分布, 很可能代表埃迪卡拉纪晚期深水环境中发育的特殊微体生物群落, 进一步厘清它们的形态和分类后, 有望成为新的埃迪卡拉纪至寒武纪界线处的指标化石。
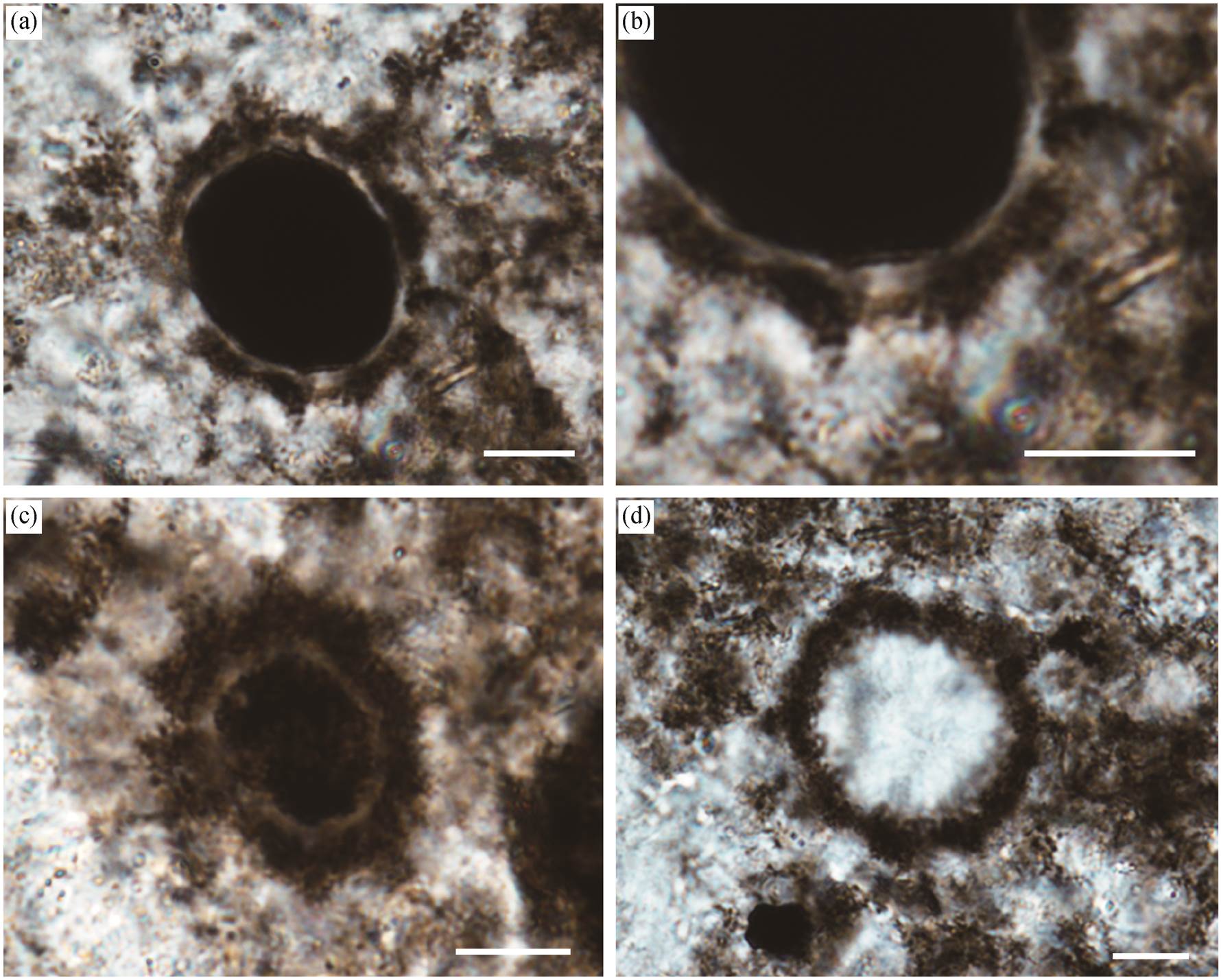
(a)~(c) 类型1, (b)为(a)突起部分放大; (d) 类型2。比例尺均代表10 μm
图5 坝黄剖面留茶坡组其他球形化石
Fig. 5 Other spherical fossils in the Liuchapo Formation at Bahuang section
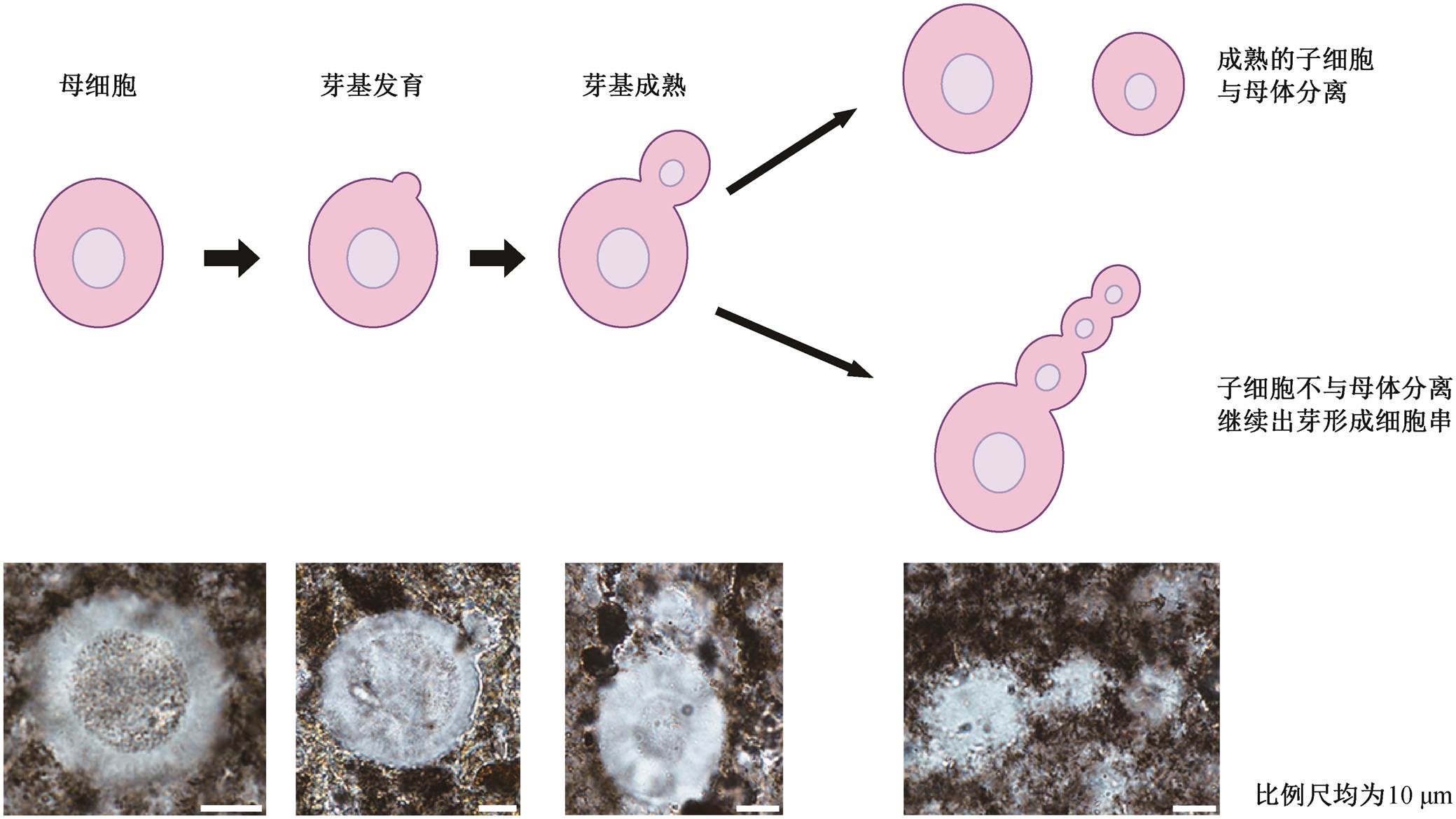
图6 球形化石Bahuangia gen. nov.与出芽生殖过程(以单细胞酵母菌为例)对比
Fig. 6 Comparison between the spherical fossil Bahuangia gen. nov. and budding process (taking unicellular yeast as an example)
Kingdom Uncertain
Phylum Uncertain
Genus Bahuangia gen. nov.
模式种 Bahuangiaconiuncta gen. et sp. nov.。
命名 属名由报道地坝黄剖面复合而成。
属征 见本属唯一种 Bahuangiaconiuncta特征。
Bahuangiaconiuncta gen. et sp. nov.
(图3(a)~(f))
(2020 “Spherical grains”, Ye等[14], Fig. 3C~D)
(2022 “Spherical fossils”, Yi等[15], Fig. 5H~M)
命名 据拉丁文coniunctus (相连的)。
正模标本 标本号 GMPKU-P-3309-1 (图 3(a)), 保存于北京大学地质博物馆。
副模标本 标本号 GMPKU-P-3310-1 (图 3(c))和 GMPKU-P-3309-2 (图 3(f)), 保存于北京大学地质博物馆。
特征 具双层结构的球形化石, 包括球形内质和外部晕圈两部分。内质为具颗粒状的球形, 位于球体中央, 其直径与球体总直径呈良好的正相关关系。外部晕圈色浅且较厚, 轮廓平滑或具小突起。球形化石以孤立或连接体的形式出现。连接体由两个以上球体以窄颈相连, 呈葫芦或链状。
测量 球体直径为 14~45μm (平均 30.4μm, SD =7.5μm, n=32), 内质直径为7~30μm (平均 18.9μm, SD=5.9μm, n=32), 二者呈较好的正相关关系(R2=0.9)。绝大多数标本中内质与球体直径比在 0.6~ 0.7 之间。
讨论 Bahuangia gen. nov.以其特殊的连接体形态与已知化石相区别, 因此本文为其建立新属。本次发现的标本中, 连接体主要为以窄颈相连的葫芦状或链状, 可与出芽生殖过程类比(见第 4 节)。基于此, Bahuangia gen. nov.可能为一类可出芽生殖的球形单细胞细菌或真菌。此外, 本研究中还观察到一例具细长丝状连接物的连接体(图 3(g))。该标本与大量 B. coniuncta gen. et sp. nov.个体共同产出, 其球体的形态和大小与 Bahuangia gen. nov.相似, 但又以连接物形态区别于 B. coniuncta gen. et sp. nov., 我们认为该标本可能为 Bahuangia gen. nov.的另一个种, 但由于仅发现一例, 暂不做进一步的划分。
除本文标本外, 前人在贵州漾头剖面留茶坡组描述的“球形颗粒”[14]以及湖南龙鼻嘴剖面留茶坡组中描述的“球形化石”[15]均表现出与 Bahuangia gen. nov.相似的特征。例如, 一些标本内部具有暗色“内核”(如文献[14]中 Fig.3C~D, 文献[15]中 Fig.5M); 它们的直径范围与 Bahuangia gen. nov.基本上重叠, 其中“球形颗粒”的直径为 8~24μm, “球形化石”的直径为 10~25μm; 在某些标本中也可观察到连接体(如文献[15]中 Fig.5H~I)。综上所述, 这些“球形颗粒”[14]以及“球形化石”[15]也应归入 Bahuangia gen. nov.。
1)在贵州省东部坝黄剖面留茶坡组的中下部, 大量产出一种具连接体的球形化石, 具球形内质及外晕圈的双层结构。在此类球形化石中, 还发现多种类型的连接体(如以短颈相连, 呈葫芦状和链状等), 这些连接体的形态可与细菌或真菌的出芽生殖类比。本文对该球形化石的形态进行了系统的描述, 并为其建立新属Bahuangia gen. nov.。
2)坝黄剖面留茶坡组中的球形化石在扬子板块埃迪卡拉纪晚期的深水相地层中分布广泛, 可能是埃迪卡拉纪晚期深水环境中的一类特殊微体生物群落。本文报道的坝黄剖面留茶坡组球形化石进一步丰富了该地区埃迪卡拉纪晚期微体生物群落的面貌, 可为区域地层对比以及地球早期生命演化的研究提供新的材料。此外, 这些球形化石具有成为埃迪卡拉纪晚期指标化石的潜力, 其细致的生物属性值得进一步研究与讨论。
致谢 感谢北京大学地球与空间科学学院孙元林教授提供宝贵意见。
参考文献
[1] Narbonne G M. The Ediacara Biota: neoproterozoic origin of animals and their ecosystems. Annual Re-view of Earth and Planetary Sciences, 2005, 33(1): 421–442
[2] 戎嘉余, 黄冰. 生物大灭绝研究三十年. 中国科学: 地球科学, 2014, 44(3): 377–404
[3] 朱茂炎, 赵方臣, 殷宗军, 等. 中国的寒武纪大爆发研究: 进展与展望. 中国科学: 地球科学, 2019, 49(10): 1455–1490
[4] 唐烽, 高林志, 尹崇玉, 等. 华南伊迪卡拉(震旦)系顶部建阶层型和界线层型新资料. 地质通报, 2015, 34(12): 2150–2162
[5] Dong L, Shen B, Lee C T A, et al. Germanium/silicon of the Ediacaran-Cambrian Laobao cherts: implica-tions for the bedded chert formation and paleoen-vironment interpretations. Geochemistry, Geophysics, Geosystems, 2015, 16(3): 751–763
[6] Jiang G, Shi X, Zhang S, et al. Stratigraphy and paleogeography of the Ediacaran Doushantuo Forma-tion (ca. 635–551 Ma) in South China. Gondwana Research, 2011, 19(4): 831–849
[7] Brasier M, Cowie J, Taylor M. Decision on the Precambrian-Cambrian boundary stratotype. Episodes, 1994, 17(1): 3–8
[8] Gradstein F, Ogg J, Smith A. A geologic time scale Cambridge: Cambridge University Press, 2004
[9] Yao J, Xiao S, Yin L, et al. Basal Cambrian mic-rofossils from the Yurtus and Xishanblaq Formations (Tarim, North-west China): systematic revision and biostratigraphic correlation of Micrhystridium-like ac-ritarchs. Palaeontology, 2005, 48(4): 687–708
[10] Dong L, Xiao S, Shen B, et al. Basal Cambrian micro-fossils from the Yangtze Gorges area (South China) and the Aksu area (Tarim Block, Northwestern China). Journal of Paleontology, 2009, 83(1): 30–44
[11] 尹崇玉. 湖北鹤峰白果坪水井沱组(早寒武世)燧 石中微化石新资料. 微体古生物学报, 1995, 12(3): 299–306, 343–344
[12] Butterfield N J. Exceptional fossil preservation and the Cambrian explosion. Integrative and Comparative Biology, 2003, 43(1): 166–177
[13] Liu P, Xiao S, Yin C, et al. Ediacaran Acanthomorphic Acritarchs and other microfossils from Chert Nodules of the Upper Doushantuo Formation in the Yangtze Gorges Area, South China. Journal of Paleontology, 2017, 88: 1–139
[14] Ye Y, Shen J, Feng Q, et al. Microbial and animal evolution in relation to redox fluctuations in a deep-water setting of South China during the Ediacaran-Cambrian transition (ca. 551–523 Ma). Palaeogeo-graphy, Palaeoclimatology, Palaeoecology, 2020, 546: 109672
[15] Yi Y, Chen F, Algeo T J, et al. Deep-water fossil assemblages from the Ediacaran-Cambrian transition of western Hunan, South China and their biostrati-graphic and evolutionary implications. Palaeogeogra-phy, Palaeoclimatology, Palaeoecology, 2022, 591: 110878
[16] 董琳, 宋伟明, 肖书海, 等. 皖南地区埃迪卡拉系皮园村组微体和宏体化石——兼论埃迪卡拉纪–寒武纪界线. 地层学杂志, 2012, 36(3): 600–610
[17] Zhu M, Zhang J, Yang A. Integrated Ediacaran (Si-nian) chronostratigraphy of South China. Palaeogeo-graphy Palaeoclimatology Palaeoecology, 2007, 254: 7–61
[18] 王鸿祯. 中国古地理图集. 北京: 地图出版社, 1985
[19] Chen D, Zhou X, Fu Y, et al. New U-Pb zircon ages of the Ediacaran-Cambrian boundary strata in South China. Terra Nova, 2015, 27(1): 62–68
[20] Budding R P, Vonk J, Shackelford T. Encyclopedia of animal cognition and behavior. Cham: Springer Inter-national Publishing, 2019: 1–3
[21] Alberts B, Bray D, Hopkin K, et al. Essential cell biology. 4th ed. New York: Garland Science, 2013
[22] Gegembauer G, Araujo L, Pereira E, et al. Serology of paracoccidioidomycosis due to paracoccidioides lutzii. PLOS Neglected Tropical Diseases, 2014, 8: e2986
[23] Cullen P J, Sprague G F Jr. The regulation of filamentous growth in yeast. Genetics, 2012, 190(1): 23–49
[24] Gimeno C J, Ljungdahl P O, Styles C A, et al. Unipolar cell divisions in the yeast S. cerevisiae lead to filamentous growth: regulation by starvation and RAS. Cell, 1992, 68(6): 1077–1090
[25] Loron C C, François C, Rainbird R H, et al. Early fungi from the Proterozoic era in Arctic Canada. Nature, 2019, 570: 232–235
Study of the Spherical Fossils from the Ediacaran-Cambrian Liuchapo Formation in Guizhou, China
Abstract Microfossils from cherts of the Liuchapo Formation at Bahuang section in eastern Guizhou Province were studied. Through thin section observation, a large number of spherical fossils were revealed, including one type with connected spheroids, and two types with dark outer wall. The former is systematically described, and a new genus Bahuangia gen. nov. is established. The connected bodies of Bahuangia are calabash- or chain-shaped, and resemble the budding unicellular fungi or bacteria in morphology. Spherical fossils similar to these Bahuang specimens are widely distributed in the late Ediacaran deep-water facies of the Yangtze Plate. Through this study, the understanding of deep-water life forms during the ECT is improved, and new materials for both regional stra-tigraphic correlation and early life evolution are provided.
Key words spherical fossils; cherts; Liuchapo Formation; Guizhou