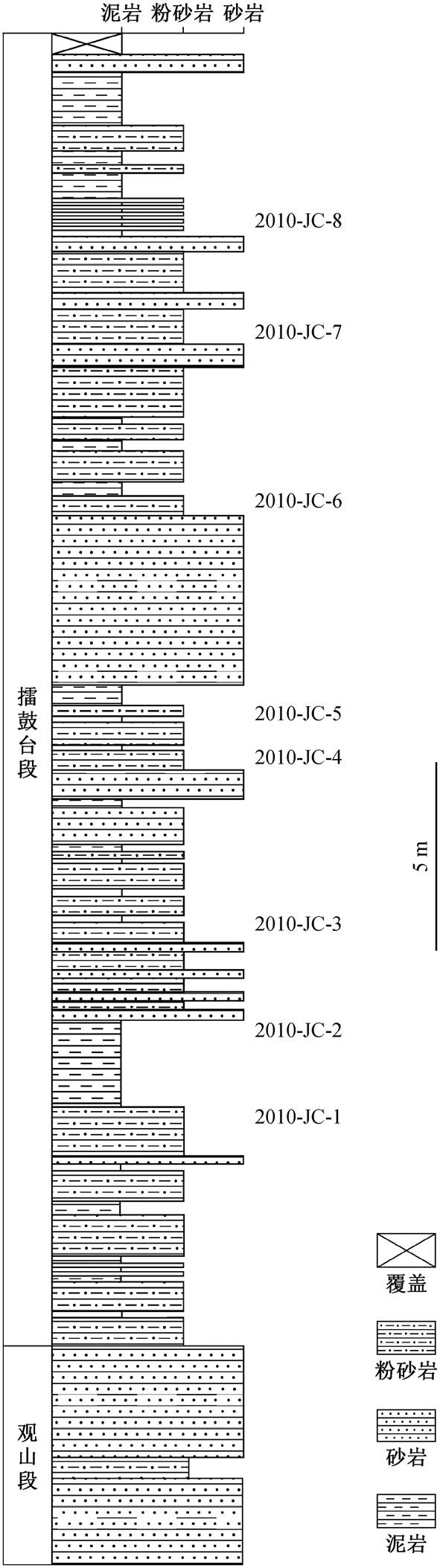
图1 箭穿剖面上泥盆统五通组地层柱状图及孢子样品采集层位
Fig. 1 Stratigraphic column of Upper Devonian Wutong Formation, at Jianchuan Section, and horizons of spore materials
北京大学学报(自然科学版) 第59卷 第2期 2023年3月
Acta Scientiarum Naturalium Universitatis Pekinensis, Vol. 59, No. 2 (Mar. 2023)
doi: 10.13209/j.0479-8023.2022.119
国家自然科学基金(42072016)资助
收稿日期: 2022‒04‒06;
修回日期: 2022‒05‒01
摘要 在安徽省广德市新杭镇箭穿剖面五通组的 8 个层位中识别出孢子 36 属 56 种及部分未定种, 并建立 1个孢子组合, 即 Apiculiretusispora hunanensis-Cymbosporites chinensis(HC)组合。与中国华南、西藏以及西欧地区孢子组合的对比结果表明, HC 组合的形成时代属于晚泥盆世法门期晚期, 层位相当于五通组擂鼓台段除顶部之外的地层。HC 组合的整体面貌与华南其他剖面的同期组合接近, 但与中国西藏及西欧地区有明显的 差别。
关键词 孢子组合; 法门期; 五通组; 擂鼓台段; 安徽
泥盆纪晚期, 维管植物迅速辐射演化, 除被子植物外的所有现生植物类群均已出现[1], 植物面貌发生明显的变化。因此, 上泥盆统与下石炭统之间的界线划分以及界线附近的古植物面貌一直是研究热点之一[2–3]。孢子化石为泥盆纪末期高分辨率的地层对比提供了依据。中国泥盆纪的多个地区地层对比主要依靠牙形石、笔石和竹节石等化石[4], 几十年来对泥盆纪孢子化石开展了大量研究[5–8], 以期进行陆相地层或受陆源输入影响较大地层间的对比。欧阳舒等[9]系统地研究了华南、准噶尔、塔里木、秦岭和西藏地区 50 多个泥盆纪孢子化石组合, 依托多门类生物地层格架, 实现这些组合的精细对比。申震[10]建立了中国泥盆纪陆生植物微体化石新数据库。然而, 关于晚泥盆世地层的顶界及划分标准, 尚存在争议。
五通组地层包括观山段(主要为石英砂岩)和擂鼓台段(石英砂岩与粉砂岩或泥岩互层), 广泛分布于安徽、江苏和浙江等地区, 具有特别丰富的植物化石。五通组常与下伏志留系茅山组平行不整合接触[11], 与上覆下石炭统金陵组连续沉积[12]。关于五通组所属时代, 目前仍有不同的认识。早期, 斯行健[13]根据植物群, 将五通组归为早石炭世。欧阳舒等[14]通过孢子分析, 认为五通组擂鼓台段顶部 Au-roraspora macra-Dibolisporites diatinctus (MD)组合带属于早石炭世。之后, 李星学等[15]结合动物、植物大化石和微体化石证据, 并对比国内外相关生物地层, 认为五通组属于晚泥盆世。高联达[8]根据长江下游五通组孢子带的研究结果, 支持五通组属于晚泥盆世; 他整合长江下游晚泥盆世法门期至早石炭世杜内期孢子, 自下而上建立 7 个组合带, 其中Rugospora flexuous-Grandispora cornuta (FC)带、Va-llatisporites pusilltes-Retispora lepidophyta(PL)带和Retispora lepidophyta-Verrucosisporites nitidus (LN)带归为法门期, 观山段的 FC 组合带属于法门期晚期, 擂鼓台段下部 PL 带和上部 LN 带属于法门期最晚期。
Wang 等[16]在安徽省广德市新杭镇发现“新杭森林”这一亚洲最古老、全球泥盆纪面积最大的化石森林, 并主要依据岩石地层序列和大化石证据与其他地层剖面进行对比, 但缺少孢粉学资料。本文的孢子样品来自该原位化石森林集中分布的五通组擂鼓台段部分地层, 籍此对相关地层的时代进行讨论。
箭穿剖面位于安徽省广德市新杭镇(剖面位置见文献[16]中图 1), 出露五通组观山段的中上部地层以及擂鼓台段部分地层, 没有展现该组的顶界。Wang 等[16]于此处报道的新杭森林为一原位石松森林, 面积至少有 25 公顷, 主体植物广德木(Guang-dedendron micrum)具有最早的根座型根系。前人对于五通组尤其是擂鼓台段的时限有争议, 而沉积岩中的锆石难以定年, 本文的孢粉学研究可为新杭森林的年代提供佐证。本次研究针对箭穿剖面擂鼓台段除顶部之外厚约 30m 的地层, 每隔 4m 间隔采样, 共得到 2010-JC-1 至 2010-JC-8 共 8 个层位的孢子样品(采样层位见图 1)。
样品处理采用《化石孢粉鉴定》标准分析方法(SY/T 5915–2018)[17], 其原理是利用化学方法溶解沉积岩中的矿物或促使化石与围岩分离, 富集包括孢粉和浮游藻类等在内的微体化石及其他各种类别的有机物。分析流程如下: 1)取样约 50g, 粉碎为粒径小于 0.5mm 的颗粒; 2)用 10%的稀盐酸浸泡, 去除钙质; 3)加满清水, 静置, 水清去水, 直至中性; 4)去水, 徐徐加入 250mL 氢氟酸, 充分搅拌, 放置 5~7 天; 5)加水, 换水 5~6 次, 直至中性; 6)用180μm 的筛子过筛, 去除大颗粒, 沉淀; 7)留 300mL左右放入离心杯, 离心去水; 8)加 150 mL浓盐酸, 置于电炉上加热约两小时, 至样品漂浮或液体澄清; 9)水洗至中性; 10)过筛法洗样, 浮选后获得全部悬浮物质。
图1 箭穿剖面上泥盆统五通组地层柱状图及孢子样品采集层位
Fig. 1 Stratigraphic column of Upper Devonian Wutong Formation, at Jianchuan Section, and horizons of spore materials
化石和有机物富集后制片, 在生物双目显微镜(型号: ZEISS Scope. A1; 目镜放大 10 倍, 物镜放大20 倍和 40 倍)下观察鉴定。孢粉化石丰富的样品至少统计 200 粒, 相对丰富的样品统计 100 粒或 4 片薄片(20mm×20mm)下的数量, 化石稀少的则统计8 片薄片。统计工作完成后, 计算其中数量大于 60粒的样品百分含量。
经显微镜下鉴定和统计, 在 8 件样品中均发现一定数量的孢子化石, 统计数量分别为 172, 179, 189, 180, 279, 70, 275 和 254 粒, 保存状况较好, 共计 36 属 56 种及部分未定种。属种和百分含量见表1。各样品中孢子面貌基本上一致, 属同一地质时期的沉积物, 可建立 Apiculiretusispora hunanensis- Cymbosporites chinensis (HC)组合。图 2 和 3 展示代表性孢子, 其组合特征如下。
表1 箭穿剖面五通组2010-JC-1至2010-JC-8各层位孢子含量统计结果(%)
Table 1 Statistical result of spore contents in horizons 2010-JC-1 to 2010-JC-8 of Wutong Formation at Jianchuan Section (%)
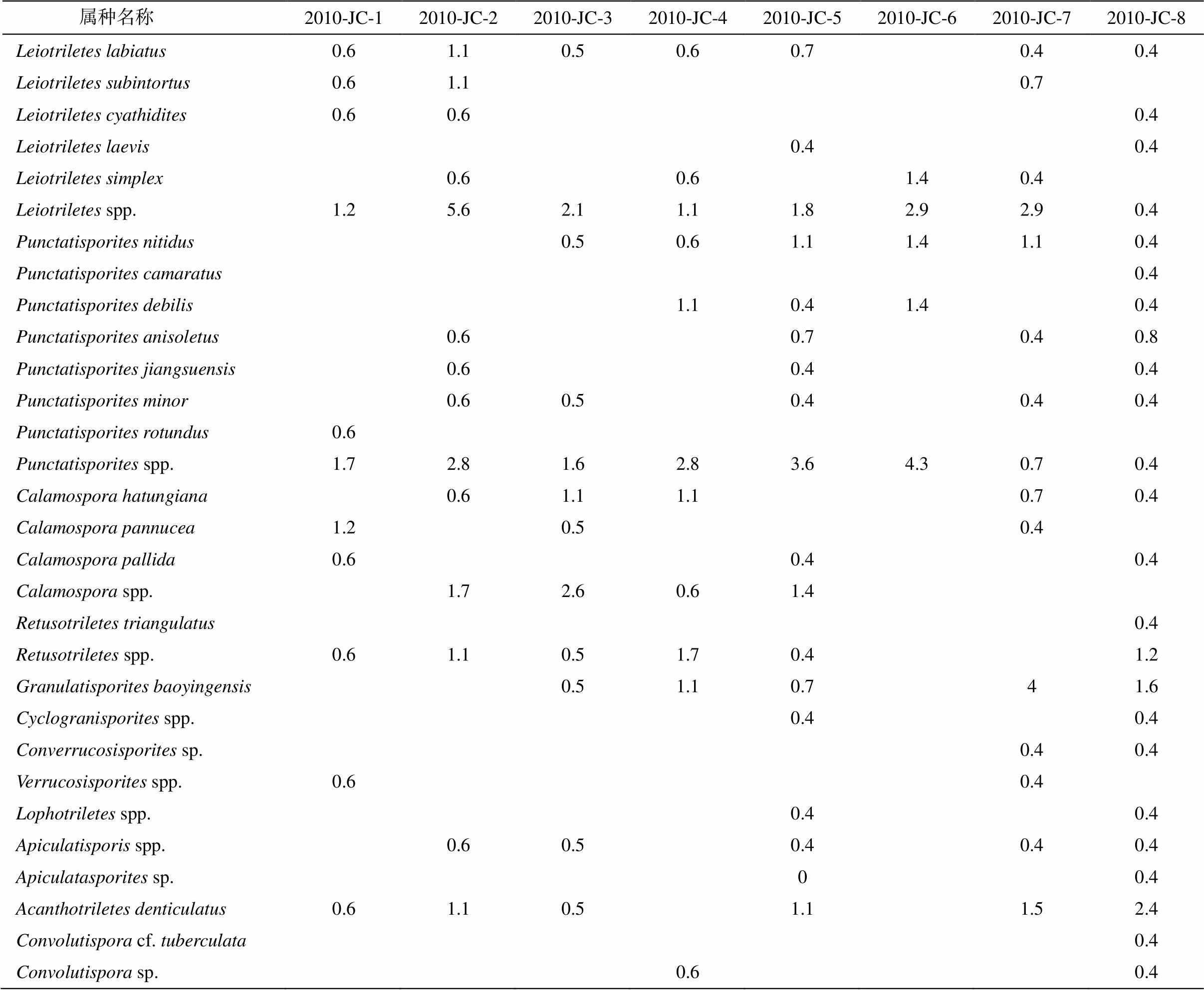
属种名称 2010-JC-12010-JC-22010-JC-32010-JC-42010-JC-52010-JC-62010-JC-72010-JC-8Leiotriletes labiatus0.61.10.50.60.70.40.4Leiotriletes subintortus0.61.10.7Leiotriletes cyathidites0.60.60.4Leiotriletes laevis0.40.4Leiotriletes simplex0.60.61.40.4Leiotriletes spp.1.25.62.11.11.82.92.90.4Punctatisporites nitidus0.50.61.11.41.10.4Punctatisporites camaratus0.4Punctatisporites debilis1.10.41.40.4Punctatisporites anisoletus0.60.70.40.8Punctatisporites jiangsuensis0.60.40.4Punctatisporites minor0.60.50.40.40.4Punctatisporites rotundus0.6Punctatisporites spp.1.72.81.62.83.64.30.70.4Calamospora hatungiana0.61.11.10.70.4Calamospora pannucea1.20.50.4Calamospora pallida0.60.40.4Calamospora spp.1.72.60.61.4Retusotriletes triangulatus0.4Retusotriletes spp.0.61.10.51.70.41.2Granulatisporites baoyingensis0.51.10.741.6Cyclogranisporites spp.0.40.4Converrucosisporites sp.0.40.4Verrucosisporites spp.0.60.4Lophotriletes spp.0.40.4Apiculatisporis spp.0.60.50.40.40.4Apiculatasporites sp.00.4Acanthotriletes denticulatus0.61.10.51.11.52.4Convolutispora cf. tuberculata0.4Convolutispora sp.0.60.4
续表
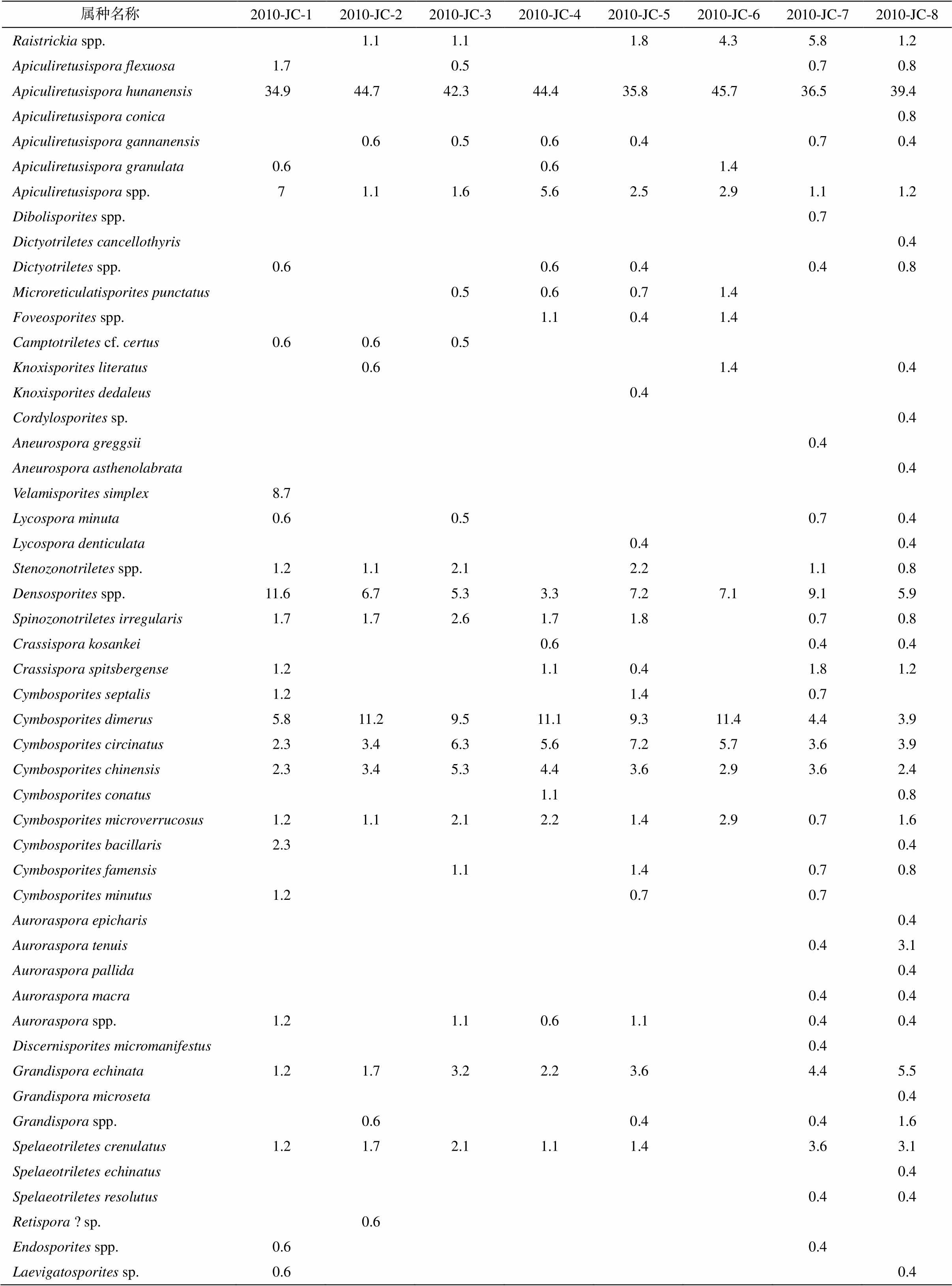
属种名称 2010-JC-12010-JC-22010-JC-32010-JC-42010-JC-52010-JC-62010-JC-72010-JC-8Raistrickia spp.1.11.11.84.35.81.2Apiculiretusispora flexuosa 1.70.50.70.8Apiculiretusispora hunanensis34.9 44.7 42.3 44.4 35.8 45.7 36.5 39.4 Apiculiretusispora conica0.8Apiculiretusispora gannanensis0.60.50.60.40.70.4Apiculiretusispora granulata0.60.61.4Apiculiretusispora spp.71.11.65.62.52.91.11.2Dibolisporites spp.0.7Dictyotriletes cancellothyris0.4Dictyotriletes spp.0.60.60.40.40.8Microreticulatisporites punctatus0.50.60.71.4Foveosporites spp.1.10.41.4Camptotriletes cf. certus0.60.60.5Knoxisporites literatus0.61.40.4Knoxisporites dedaleus0.4Cordylosporites sp.0.4Aneurospora greggsii0.4Aneurospora asthenolabrata0.4Velamisporites simplex8.7Lycospora minuta0.60.50.70.4Lycospora denticulata0.40.4Stenozonotriletes spp.1.21.12.12.21.10.8Densosporites spp.11.6 6.75.33.37.27.19.15.9Spinozonotriletes irregularis 1.71.72.61.71.80.70.8Crassispora kosankei0.60.40.4Crassispora spitsbergense1.21.10.41.81.2Cymbosporites septalis1.21.40.7Cymbosporites dimerus5.811.29.511.19.311.44.43.9Cymbosporites circinatus2.33.46.35.67.25.73.63.9Cymbosporites chinensis2.33.45.34.43.62.93.62.4Cymbosporites conatus1.10.8Cymbosporites microverrucosus1.21.12.12.21.42.90.71.6Cymbosporites bacillaris2.30.4Cymbosporites famensis1.11.40.70.8Cymbosporites minutus1.20.70.7Auroraspora epicharis0.4Auroraspora tenuis0.43.1Auroraspora pallida0.4Auroraspora macra0.40.4Auroraspora spp.1.21.10.61.10.40.4Discernisporites micromanifestus0.4Grandispora echinata1.21.73.22.23.64.45.5Grandispora microseta0.4Grandispora spp.0.60.40.41.6Spelaeotriletes crenulatus1.21.72.11.11.43.63.1Spelaeotriletes echinatus0.4Spelaeotriletes resolutus0.40.4Retispora ? sp.0.6Endosporites spp.0.60.4Laevigatosporites sp.0.60.4

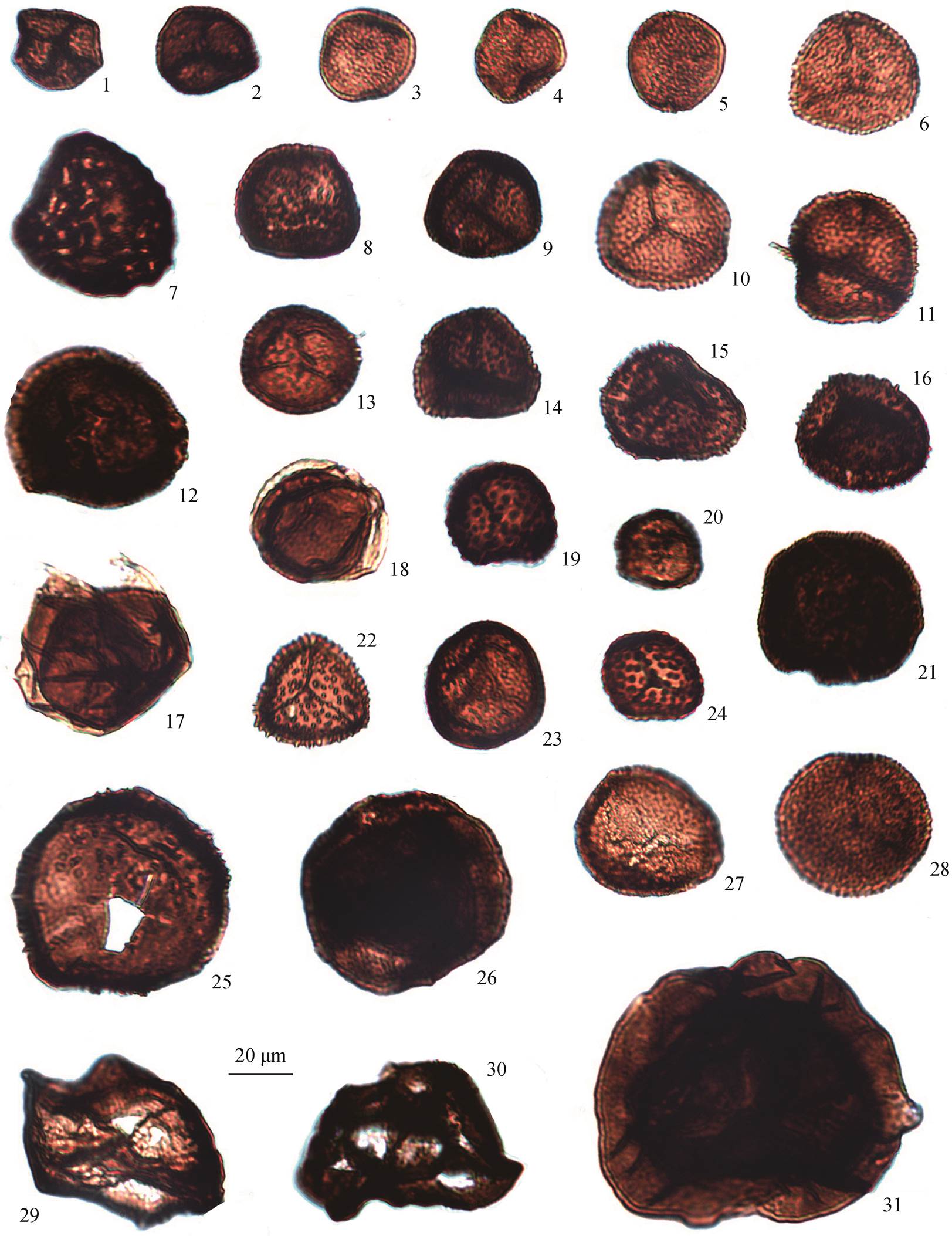
1: Punctatisporites nitidus Hoffmeister, Staplin and Malloy, 1955, No. 2010-JC-8; 2: Punctatisporites camaratus Ouyang and Chen, 1987, No. 2010-JC-8; 3: Punctatisporites debilis Hacquebard, 1957, No. 2010-JC-1; 4: Leiotriletes cyathidites Zhou, 1980, No. 2010-JC-7; 5: Leiotriletes sp., No. 2010-JC-8; 6 和 15: Raistrickia sp., No. 2010-JC-8; 7: Leiotriletes labiatus Ouyang and Chen, 1987, No. 2010-JC-1; 8: Converru-cosisporites sp., No. 2010-JC-2; 9: Leiotriletes subintortus (Waltz) Ischenko, 1952 var. rotundatus Waltz, 1941, No. 2010-JC-1; 10: Dictyotri-letes sp., No. 2010-JC-7; 11: Punctatisporites anisoletus Ouyang and Chen, 1987, No. 2010-JC-8; 12: Retusotriletes sp., No. 2010-JC-8; 13 和14: Cyclogranisporites baoyingensis Ouyang and Chen, 1987, No. 2010-JC-8; 16: Aneurospora greggsii (Mcgregor) Streel, 1974, No. 2010-JC-8; 17: Lycospora denticulata Bharadwaj, 1957, No. 2010-JC-8; 18: Microreticulatisporites punctatus Knox, 1950, No. 2010-JC-3; 19: Calamos-pora pannucea Richardson, 1965, No. 2010-JC-3; 20: Calamospora hatungiana Schopf, 1944, No. 2010-JC-8。标本均保存于北京大学地球与空间科学学院地质学系
图2 安徽新杭五通组(擂鼓台段除顶部之外的地层)代表性孢子(第一部分)
Fig. 2 Representative spores from the Wutong Formation (Leigutai Member except the uppermost port) in Xinhang, Anhui (part one)
1 和 2: Apiculiretusispora flexuosa Hou, 1982, No. 2010-JC-2, 2010-JC-1; 3~5: Apiculiretusispora hunanensis (Hou) Ouyang and Chen, 1987, No. 2010-JC-8; 6: Apiculiretusispora conica Lu and Ouyang, 1976, No. 2010-JC-8; 7: Camptotriletes cf. certus Venkat. et Bharad., 1962, No. 2010-JC-3; 8和9: Cymbosporites microverrucosus Bharadwaj, Tiwari and Venkatachala, 1971, No. 2010-JC-7, 2010-JC-1; 10: Cymbosporites chinensis Ouyang and Chen, 1987, No. 2010-JC-8; 11: Cymbosporites conatus Bharadwaj et al., 1971, No. 2010-JC-8; 12和21: Densosporites sp., No. 2010-JC-8; 13和14: Cymbosporites circinatus Ouyang and Chen, 1987, No. 2010-JC-8, 2010-JC-2; 15和16: Grandispora echinata Hacquebard, 1957, No. 2010-JC-7; 17和18: Auroraspora tenuis Ouyang and Chen, 1987, No. 2010-JC-8; 19和24: Cymbosporites dimerus Ouyang and Chen, 1987, No. 2010-JC-2, 2010-JC-8; 20: Lycospora minuta (Ischenko) Somers, 1972, No. 2010-JC-8; 22: Lycospora sp., 2010-JC-8; 23: Cymbosporites septalis (Jush.) var. minor (Kedo), 1994, No. 2010-JC-8; 25: Spinozonotriletes irregularis Liu et Ouyang, 2020, No. 2010-JC-4; 26: Auroraspora epicharis Zhu, 1999, No. 2010-JC-1; 27: Aneurospora asthenolabrata (Hou) Lu, 1994, No. 2010-JC-8; 28: Spelaeotriletes crenulatus (Playford) Higgs, Claytan and Keegan, No. 2010-JC-8; 29: Knoxisporites literatus (Waltz) Playford, 1963, No. 2010-JC-2; 30: Knoxisporites dedaleus (Naumova) Lu, 1994, No. 2010-JC-8; 31: Velamisporites simplex Ouyang and Chen, 1987, No. 2010-JC-1。标 本均保存于北京大学地球与空间科学学院地质学系
图3 安徽新杭五通组(擂鼓台段除顶部之外的地层)代表性孢子(第二部分)
Fig. 3 Representative spores from the Wutong Formation (Leigutai Member except the uppermost port) in Xinhang, Anhui (part two)
1)无环三缝孢类占优势, 其含量为 56.63%~70.0%, 平均 62.59%; 具环/腔三缝孢类次之, 含量为 30.0%~43.37%, 平均 37.29%; 单缝孢类最少, 含量为 0~0.58%, 平均 0.12%; 未见任何可靠的裸子植物花粉。
2)无环三缝孢类中, Apiculiretusispora占绝大多数, 含量为 38.71%~51.11%, 平均 44.60%, 主要为 A. hunanensis; 其次为A. flexuosa, A. conica, A. gannanensis以及 A. granulata等。Punctatisporites和 Leiotriletes 含量较高。Punctatisporites 占 2.33%~7.14%, 平均 4.15%, 主要为 Punctatisporites nitidus, P. debilis, P. anisoletus 和 P. minor 等。Leiotriletes占 1.57%~8.94%, 平均 3.73%, 主要为 Leiotriletes labiatus, L. subintortus 和 L. simplex等。其余分子(如 Retusotriletes, Granulatisporites baoyingensis, Api-culatisporis, Acanthotriletes denticulatus, Raistrickia, Dibolisporites echinaceus, Dictyotriletes, Microreti-culatisporites punctatus, Camptotriletes cf. certus,Knoxisporites literatus, K. dedaleus, Aneurospora gre-ggsii 和 Velamisporites simplex 等)少见。
3)具环/腔三缝孢类中, Cymbosporites 含量最高, 占 13.78%~25.09%, 平均 20.04%, 主要为 C. di-merus, C. circinatus 和 C. chinensis; 其次为 C. septa-lis, C. conatus, C. microverrucosus 和 C. famensis等。Densosporites 和 Grandispora/Spelaeotriletes 也有一定的含量。Densosporites 占 3.33%~11.63%, 平均7.03%。Grandispora/Spelaeotriletes 占 0~11.42%, 平均 5.05%, 主要为 Grandispora echinata和 Spelaeo-triletes echinatus 等。其余分子(如 Lycospora minu-ta, L. denticulate, Stenozonotriletes, Spinozonotrile-tes irregularis, Crassispora spitsbergense, Auroras-pora tenuis, Auroraspora macra, Discernisporites mi-cromanifestus 和Endosporites 等)少见。
4)单缝孢类贫乏, 仅 Laevigatosporites 有零星的分布。
根据孢子属种和数量分析, 本文中 HC 组合最突出的特征是具纹饰弓形脊的 Apiculiretusispora 以及外壁呈明显栎状加厚的 Cymbosporites 的高含量。其中, Apiculiretusispora hunanensis 最早发现于湖南锡矿山组马牯脑段顶部、欧家冲组和邵东组下部, 地质时代为法门期晚期[18], 此后在江苏、浙江、西藏和新疆塔里木等地的法门期晚期孢子组合中也有报道[5,7,18‒21], 该分子目前仅发现于晚泥盆世法门期晚期地层中。Cymbosporites chinensis 和 C. circina-tus 为华南地区晚泥盆世晚期的常见分子, 如江苏句容五通组擂鼓台段下部[5]、湖南锡矿山欧家冲组和邵东组[18]、江西全南三门滩组[22]和西藏聂拉木波曲群上部[19]等。Acanthotriletes denticulatus 为一全球广布但限于晚泥盆世晚期的孢子种, 首见于俄罗斯地台晚泥盆世[23], 在我国见于湖南锡矿山和界岭邵东组上部[18]、江苏南京龙潭五通组观山段‒擂鼓台段下部[6]和江西全南三门滩组[22]等法门期晚期地层中。
其他主要或仅限于晚泥盆世晚期的分子还有Apiculiretusispora flexuosa, A. gannanensis, A. gra-nulate, Leiotriletes labiatus, Punctatisporites minor, Granulatisporites baoyingensis, Cymbosporites dim-erus, C. famensis 和 C. minutus等。法门期晚期‒早石炭世的过渡分子有 Grandispora echinata, Discer-nisporites micromanifestus, Knoxisporites literatus 和Auroraspora macra 等。个别分子呈现明显的石炭纪色彩, 如 Lycospora minuta, L. denticulata 和 Cras-sispora kosankei 等。
综上所述, 本文的 HC 组合中出现极少数石炭纪分子, 且缺乏早‒中泥盆世常见的 Rhabdosporites, Calyptosporites, Emphanisporites, Chelinospora, An-cyrospora以及 Archaeoperisaccus 等。因此, 我们认为当前组合代表的地层时代应为晚泥盆世法门期晚期。
我国泥盆纪孢子化石的研究较为深入, 已建立相对完整的孢子组合带, 其中晚泥盆世晚期的孢子记录主要分布于江苏、浙江、湖南、江西、西藏、新疆、贵州和云南等地[5,7,18–22,24–27]。
目前对安徽地区晚泥盆世孢子化石的研究较少。高联达等[28]报道了皖南休宁和歙县地区溪口群浅变质岩中的 4 个剖面的孢子组合, 所属时代一致, 均为晚泥盆世法门期, 但不是最晚期。这些组合中的孢子以具膜环、瘤面和具环瘤面类为主, 如Grandispora famenensis, G. inculta, G. saurota, G.gracilis, Verrucosisporites nitidus, Lophotriletes me-dia, L. gibberulus 和 L. proscurrus 等。这些孢子都没有在本文的 HC 组合中出现, 而本文 HC 组合中的一些优势分子在皖南浅变质岩孢子组合[28]中也未见及。由于研究区不同, 皖南浅变质岩孢子组合与本文的 HC 组合的主要孢子类型存在明显的差异, 可对比性不大。
本文的 HC 组合与江苏句容包 1 井晚五通组擂鼓台段下部 Retispora lepidophyta var. minor-Apicu-liretusispora hunanensis-Cymbosporites spp.组合[5]的孢子面貌十分相似, 地质时代(法门期晚期)相当。这两个组合均以丰富的 Apiculiretusispora和 Cym-bosporites 为主要特征, 含大量相同的种, 如 Apicu-liretusispora hunanensis, A. granulate, Cymbosporitesdimerus, C. circinatus, C. chinensis, C. minutus, Kno-xisporites literatus, Leiotriletes laevis, L. labiatus, L.subintortus, L. simplex, Punctatisporites camaratus, P. rotundus, P. anisoletus, P. jiangsuensis, Calamo-spora pallida, Retusotriletes triangulates, Velamispo-rites simplex, Lycospora denticulate 和 Grandisporaechinata 等。唯一的不同之处是, 本文的 HC 组合中未发现包 1 井下部组合的主要分子 Retispora lepido-phyta var. minor。与包 1 井下部组合主要成分类似、地质时代相当的组合还见于江苏宝应地区应 2 井上部组合[14]。
江苏南京龙潭五通组擂鼓台段中下部 Retuso-triletes-Cymbosporites (RC)组合[6], 地质时代为晚泥盆世法门期晚期。该组合以 Cymbosporites的大量出现为主要特征, 含量可达 26.8%, 有 C. conatus, C. zonalis, C. famensis, C. septalis, C. microverruco-sus和 C. dimerus 等种, 还有一定数量的 Punctati-sporites和 Retusotriletes, 其他重要分子有 Apiculi-retusispora flexuosa, A. gannanensis, A. granulate,Leiotriletes labiatus, Punctatisporites rotundus, Gra-nulatisporites baoyingensis, Acanthotriletes denticu-latus, Discernisporites micromanifestus, Knoxispori-tes literatus和 Grandispora echinata 等。这些分子基本上都出现在本文的 HC 组合中, 两个组合的相似性较高。
何圣策等[7]将浙江富阳西湖组划分为两个孢子组合, 下部为 Retispora lepidophyta-Apiculiretusis-pora hunanensis (LH)组合, 属晚泥盆世法门期晚期。主要分子为 Apiculiretusispora hunanensis, A. nitida, Retusotriletes, Knoxisporites literatus 和 Auroraspora cf. poljessica, 其次为 Punctatisporites, Leiotriletes, Lophotriletes uncutas, Densosporites xihuanensis,Cymbosporites, Grandispora, Auroraspora macra 和Retispora lepidophyta 等。虽然该组合的分异度低于本文的 HC 组合, 且出现本文 HC 组合中缺少的 Re-tispora lepidophyta, 但从总体面貌来看, 两个组合的主要孢子属种相近。
湖南锡矿山地区欧家冲剖面马牯脑段、欧家冲组和邵东组下部的第一孢子组合带, 即 Retispora lepidophyta-Granulatisporites (Apiculiretusispora)hu-nanensis 组合带[18], 地质时代属于晚泥盆世法门期晚期。该组合属的种多样性相对贫乏, Apiculiretu-sispora hunanensis, Cymbosporites parvibasilaris和Retispora lepidophyta 占据优势, 其他主要分子有 A.flexuosa, Acanthotriletes denticulatus, Knoxisporitesliteratus, Discernisporites micromanifestus, Leiotrile-tes, Punctatisporites, Retusotriletes incohatus, Lopho-triletes, Grandispora和 Spelaeotriletes 等。这些属种多见于本文的 HC 组合, 表明二者层位大致相当。
文子才等[22]报道了江西全南小慕泥盆纪–石炭纪界线附近 3 个孢子组合带, 其中法门期晚期翻下组 Leiotriletes macrothelis-Grandispora xiaomuensis (MX)组合与本文中 HC 组合关系较密切。共同出现的重要分子有 Cymbosporites circinatus, Grandispo-ra echinata, Aneurospora asthenolabrata, A. greggsii, Apiculiretusispora gannanensis, Discernisporites mic-romanifestus, Auroraspora macra, Leiotriletes laevis, L. simplex, Punctatisporites minor, P.jiangsuensis 和Acanthotriletes denticulatus 等, 但翻下组中的带分子 Leiotriletes macrothelis 和 Grandispora xiaomuensis却未在本文 HC 组合中发现, 其原因可能是二者沉积环境不同, 翻下组属于近岸海相碎屑岩沉积, 而五通组擂鼓台段除顶部之外的地层为非海相沉积。
高联达[29]在西藏珠峰地区建立的 Retispora le-pidophyta-Hymenozonotriletes explanatus (LE)带, 地质时代属于法门期晚期, 与本文的 HC 组合相比, 共同属种不多。Retispora lepidophyta在西藏珠峰地区上泥盆统章东组中数量丰富(占总量的 20.0%~30.0%或更多), 是其中典型属种, 并且常见 Hymeno-zonotriletes explanatus和 Vallatisporites pusillites伴随出现。我国西藏地区LE带的总体面貌与西欧地区LE带的吻合度很高, 相比之下, 本文中HC 组合带的地方性特征更明显, 如 Apiculiretusispora huna-nensis, A. gannanensis, Auroraspora macra和 Cym-bosporites chinensis等属种通常被认为是华南法门期晚期的典型孢子种类[9,18,30]。
目前, 西欧地区泥盆纪–石炭纪孢子的研究程度比较高, 将晚泥盆世法门期自下而上分为4个孢子组合带: Rugospora versabilis-Grandispora uncata (VU)带、Retispora lepidophyta-Knoxisporites litera-tus(LL)带、Retispora lepidophyta-Hymenozonotriletes explanatus(LE)以及Retispora lepidophyta-Verrucosi-sporites nitidus(LN)带, 已在全球范围的法门期地层内得到广泛的识别[31‒32]。比利时狄南盆地和纳谬尔盆地晚泥盆世法门期晚期LE孢子组合带[33‒34]以含Retispora lepidophyta和 Vallatisporites pusillites为主要特征, 这两个种也在前苏联 Prypyat 盆地[35‒36]以及美国俄亥俄州和蒙大拿州[37‒38]晚泥盆世(法门期)地层中大量发现。其中, Retispora lepidophyta是一个全球性的种, 在法门期晚期大量出现, 在泥盆纪–石炭纪界线处消失, 目前视此种为晚泥盆世晚期的标准分子。虽然这种孢子在华南出现, 但数量很少, 如在江苏句容擂鼓台组下部占 0.26%[5], 在南京龙潭擂鼓台段中下部占1.2%[6], 也出现在南黄海擂鼓台段中下部[39], 但远不如我国西藏和西欧地区那样丰富(有时可占组合带内孢子总量的半数以上), 且在形态上也不如西欧报道的那样典型(穴或网不明显), 在鉴定方面尚存分歧。
综上所述, 本文的HC组合属于晚泥盆世法门期晚期, 层位对应五通组擂鼓台段除顶部之外的地层。本组合与江苏、浙江、江西和湖南等地同期的孢子组合特征接近, 似乎更具区域性或地方性, 而与我国西藏及西欧等地区明显不同。造成该孢子组合差异的原因除沉积环境外, 可能还与不同的植物面貌有关。
本文在安徽省新杭镇箭穿剖面五通组擂鼓台段中下部识别出孢子36属56种及部分未定种, 建立一个孢子组合: Apiculiretusispora hunanensis-Cym-bosporites chinensis (HC)组合, 属法门期晚期。
HC组合与江苏句容、宝应及南京龙潭等地的五通组中下部、浙西富阳的西湖组、湖南中部的马牯脑段、欧家冲组和邵东组下部、江西全南的翻下组同期孢子组合特征相似度高, 时代相当, 但与我国西藏和西欧地区的LE带有明显差异。
致谢 研究工作得到中国科学院南京地质古生物研究所黄璞博士的诸多帮助, 谨表谢忱。
参考文献
[1] 郝守刚, 王德明, 王祺. 陆生植物的起源和维管植物的早期演化. 北京大学学报(自然科学版), 2002, 38(2): 286–293
[2] Xiong C H, Wang D M, Wang Q, et al. Diversity dynamics of Silurian–Early Carboniferous land plants in South China. PLOS ONE, 2013, 8(9): e75706
[3] Cascales-Miñana B. Apparent changed in the Ordovi-cian–Mississippian plant diversity. Review of Palaeo-botany and Palynology, 2016, 227: 19–27
[4] 郄文昆, 马学平, 徐洪河, 等. 中国泥盆纪综合地层和时间框架. 中国科学: 地球科学, 2019, 49(1): 115–138
[5] 欧阳舒, 陈永祥. 江苏句容泥盆–石炭系孢子组合并讨论五通群的时代问题. 中国科学院南京地质古生物研究所集刊, 1987(23): 1–92
[6] 卢礼昌. 南京龙潭地区五通组孢子组合及其地质时代. 微体古生物学报, 1994, 11(2): 153–199
[7] 何圣策, 欧阳舒. 浙西富阳西湖组泥盆–石炭系过渡层孢子组合. 古生物学报, 1993, 32(1): 31–48
[8] 高联达. 长江下游地区晚泥盆世至早石炭世孢子带和泥盆系–石炭系界线. 地球学报, 2015, 36(1): 7–21
[9] 欧阳舒, 卢礼昌, 朱怀诚, 等. 中国晚古生代孢粉化石. 合肥: 中国科学技术大学出版社, 2017
[10] 申震. 泥盆纪孢子和疑源类的时空分布及其意义: 以西准噶尔、华南和数据库为例[D]. 武汉: 中国地质大学(武汉), 2020
[11] 戎嘉余, 陈旭, 王怿. 志留系//中国科学院南京地质古生物研究所. 中国地层研究二十年(1979—1999). 合肥: 中国科学技术大学出版社, 2000: 73–95
[12] 侯明金, 齐敦伦. 安徽巢湖地区泥盆纪–石炭纪界线地层的新认识. 地层学杂志, 2006, 30(2): 157–170
[13] 斯行健. 江苏乌桐系Lepidodendropsis hirmeri Lutz的发现与讨论. 古生物学报, 1956, 4(1): 45–52
[14] 欧阳舒, 陈永祥. 江苏中部宝应地区晚泥盆世–早石炭世孢子组合. 微体古生物学报, 1987, 4(2): 195–214
[15] 李星学, 蔡重阳, 欧阳舒. 长江下游五通组研究的新进展. 中国地质科学院院报, 1984(2): 119–133
[16] Wang D M, Qin M, Liu L, et al. The most extensive Devonian fossil forest with small lycopsid trees bea-ring the earliest Stigmarian roots. Current Biology, 2019, 29: 2604–2615
[17] SY/T 5915—2018. 化石孢粉鉴定[S/OL]. (2018–10–29)[2022–01–10].https://www.cssn.net.cn/cssn/produ-ctDetail/41b1788b5a34f7bc8218286e6f823b39
[18] 侯静鹏. 湘中锡矿山地区泥盆–石炭系过渡层的孢子组合. 中国地质科学院地质研究所文集, 1982(5): 81–95
[19] 高联达. 西藏聂拉木晚泥盆世孢子的发现及其地层意义. 青藏高原地质文集, 1983(1): 183–218, 255–262
[20] 朱怀诚, 赵治信. 塔里木盆地泥盆–石炭系孢粉研究新进展. 新疆石油地质, 1999(3): 74–77
[21] Liu F, Kerp H, Peng H P, et al. Palynostratigraphy of the Devonian-Carboniferous transition in the Tulong section in South Tibet: a Hangenberg event sequence analogue in the Himalaya-Tethys zone. Palaeogeo-graphy, Palaeoclimatology, Palaeoecology, 2019, 531: 108704
[22] 文子才, 卢礼昌. 江西全南小慕泥盆–石炭系孢子组合及其地层意义. 古生物学报, 1993, 32(3): 303–331, 405–408
[23] Naumova S N. Spore-pollen assemblages of the Upper Devonian of the Russian Platform and their stratigra-phic significance. Transactions of Institute of Geolo-gical Sciences, Academy of Sciences of USSR, 1953, 143(60): 1–204
[24] 卢礼昌. 关于我国“Retispora lepidophyta”的再研究. 微体古生物学报, 1994, 11(4): 469–478
[25] 高联达. 贵州东南部晚泥盆世至早石炭世孢子带和泥盆–石炭系界线. 贵州地质, 1991(1): 59–71
[26] 朱怀诚. 塔里木盆地北部东河塘组孢子化石. 古生物学报, 1999, 38(3): 69–87
[27] Yang W. Stratigraphic and phytogeographic palyno-logy of late Paleozoic sediments in western Yunnan, China. Science Reports of Niigata University (Geo-logy), 1999, 14: 15–99
[28] 高联达, 王涛, 王宗起, 等. 安徽南部地区浅变质地层孢子化石的发现及其地质意义. 地质通报, 2014, 33(9): 1267–1273
[29] 高联达. 西藏聂拉木晚泥盆世至早石炭世孢子带和泥盆–石炭系界线//喜马拉雅岩石圈构造演化古生物论文集. 北京: 地质出版社, 1988: 181–247
[30] Ouyang S, Chen Y. Palynology of Devonian–Carboni-ferous transition sequences of Jiangsu, E. China. Pa-laeontologia Cathayana, 1989, 4(4): 439–473
[31] Clayton G, Coquel R, Doubinger J, et al. Carboni-ferous miospores of western Europe: illustration and zonation. Mededelingen Rijks Geologische Dienst, 1977, 29: 1–71
[32] Marshall J E A. A terrestrial Devonian–Carboniferous boundary section in East Greenland. Palaeobiodi-versity and Palaeoenvironments, 2021, 101: 541–559
[33] Streel M. Distribution stratigraphique et Géographi-qued’ Hymenozonotriletes lepidophytus Kedod’ Hy-menozonotriletes pusillites — Kedo et des assembla-ges tournaisiens (Synthèse du projet B. Congrèset Collowues Université de Liège, 1970, 55: 121–147
[34] Maziane N, Higgs K T, Streel M. Revision of the late Famennian miospore zonation scheme in eastern Bel-gium. Journal of Micropalaeontology, 1999, 18(1): 17–25
[35] Kedo G I. Spores from the supra salt Devonian deposits of the Pripyat Depression and their strati-graphic significance. Paleontologie and Stratigraphy of Russia, Akademiya Nauk Belorussian Soviet Socialist Republics, Minsk, 1957, 2: 3–43
[36] Kedo G I. Spores of the Tournaisian stage of the Pripyat Depression and their Stratigraphical signifi-cance. Paleontologiya i Stratigrafiya Belorussian So-viet Socialist Republics, Minsk, 1963, 4: 3–127
[37] Winslow M R. Plant spores and other microfossils from Upper Devonian and Lower Mississippian rocks of Ohio. United States Geological Survey, Profes-sional Paper, 1962, 364: 1–27
[38] Sandberg C A, Streel M, Scott R A. Comparison between conodont zonation and spore assemblages at the Devonian–Carboniferous boundary in the western and central United States and in Europe. Compte Ren-du, Septieme Congres International de Stratigraphie et de Geologie du Carbonifere, Krefeld, 1971, 1: 179–203
[39] Guo X W, Zhang X H, Cai L X, et al. Late Devonian–Early Carboniferous palynology of the CSDP-2 Borehole in the southern Yellow Sea, China. Palaeo-world, 2022, 31: 258–276
Spore Assemblage from the Upper Devonian Wutong Formation in Xinhang, Anhui
Abstract Spores of 36 genera, 56 species and some indeterminate species are recognized from 8 layers of Wutong Formation at Jianchuan Section, Xinhang County, Guangde City, Anhui Province. A spore assemblage can be established, that is Apiculiretusispora hunanensis-Cymbosporites chinensis (HC) Assemblage. By comparing the main components of HC Assemblage with spore assemblages in South China, Tibet of China and Western Europe, HC Assemblage is assigned to the late Famennian of the Late Devonian and corresponds to the strata except the uppermost part of the Leigutai Member of the Wutong Formation. The overall characteristics of HC Assemblage are close to contemporaneous assemblages of South China but different from those of Tibet of China and Western Europe.
Key words spore assemblages; Famennian; Wutong Formation; Leigutai Member; Anhui