红树植物耐盐–耐淹性的荟萃分析及其应用对策
江鎞倩 李瑞利† 沈小雪 张志 张月琪
北京大学深圳研究生院环境与能源学院, 深圳 518055; †通信作者, E-mail: liruili@pkusz.edu.cn
摘要 基于盐度和潮汐淹水对红树植物筛选和定植的重要作用, 选择 5 种中国造林工程中常用的红树植物, 采用文献检索和荟萃分析方法, 探究其耐盐–耐淹性。结果表明: 白骨壤(Avicennia marina)的耐盐–耐淹性最强, 能够生长于高盐度(40‰)和长时间淹水(16h/d)环境中; 秋茄(Kandelia obovata)和桐花树(Aegiceras corniculatum)具有较强的耐盐–耐淹性, 能够生长于中盐度(30‰)和较长时间淹水(12h/d)环境中; 木榄(Bruguiera gymnorrhiza)和无瓣海桑(Sonneratia apetala)的耐盐性较低, 能够生长于低盐度(20‰)环境中, 前者可种植在较长时间淹水(12h/d)区域; 超过单一盐度或淹水胁迫耐受限度时, 复合胁迫会降低红树植物的耐受性; 在实际造林应用中, 白骨壤、秋茄和桐花树具有较强的耐盐–耐淹性, 适合中国南部沿海的红树林宜林地。最后, 提出相应的工程应用对策, 可为中国红树林的精准修复提供科学依据。
关键词 红树植物; 耐盐; 耐淹; 人工造林; 生态修复
红树林是生长在热带和亚热带海岸潮间带的木本植物群落, 具有重要的生态、社会和经济价值[1]。作为地球碳含量最大的生态系统之一[2], 每公顷红树林平均含碳 1023 吨[3], 通过提供食物、繁衍和育苗的栖息地吸引鱼虾[4], 可以有效地抵御极端洪水事件, 保护海岸线[5], 同时还具有促淤造陆的生态功能[6–7]。由于人类活动和气候变化, 全球红树林面积正在大量减少。2000—2016 年间, 全球红树林面积因土地利用变化减少 62%[8]。红树林锐减的主要原因是其转化为水产养殖业和农业用地[9], 其次为道路建设、非法伐木、城市化建设以及旅游开发等[10]。此外, 气候变化也会改变潮汐沼泽的结构和功能, 如降雨增加会导致淡水流入和海平面上升, 从而改变红树林生境[11–12]。我国的红树林主要分布在广东、广西和海南等地[13], 是保护中国濒危生物和跨国鸟类的重要区域[14]。历史上, 我国红树林面积曾达 25 万 hm2 [15]。1973—2000 年间, 红树林面积从 48801hm2 降至 18702hm2, 之后得益于对剩余红树林的保护和恢复行动, 其面积以每年 1.8%的速度增长, 2015 年时增至 22419hm2。截至 2019 年, 中国 67%的红树林被列入保护区, 得到较好的保护[16–17]。虽然中国红树林的恢复初见成效, 但还有艰巨的任务需要完成。
2020 年, 我国首个红树林保护修复专项行动计划出台, 明确提出对浙江、福建、广东、广西和海南现有红树林实施全面保护和修复。由于自然界存在一些红树生长限制因素, 红树林恢复项目往往完全失败或达不到目标, 人工红树林的死亡率很高, 其中一个重要原因就是造林物种与种植环境不匹配[18–19]。由于不了解红树植物对种植环境的耐受程度[20], 因此不能评估造林物种合适的水文和生态条件[21]。盐度和淹水是红树植物生长的重要环境因素。盐度会影响红树植物的生长、离子水平、组织结构、热值变化、抗氧化酶活性以及相容性物质合成等[22]。盐分的增加会严重地抑制土壤微生物的养分循环和释放, 导致红树林土壤因养分贫乏而退化, 进而破坏由潮汐主导的盐度梯度, 对盐度敏感的物种将不能适应环境的变化[23]。由于红树植物生长在海岸潮间带, 所以大部分时间处在潮水淹没状态中, 有的甚至处于没顶淹水状态[24–25]。红树林在潮间带和河口地区的分布差异, 是浮力对其繁殖体的分选以及繁殖体在潮间带建立的能力差异造成的[26]。由于海平面上升和人为压力日益增加, 潮汐淹水可能造成某些红树物种消亡, 从而威胁红树林的分布和多样性[27]。同时, 潮汐和盐度的交互作用对红树林的影响不亚于单一因素。因此, 在选择造林物种和造林区域时, 应该着重考虑当地环境的盐度和淹水情况。
基于上述背景, 本研究选取秋茄、白骨壤、木榄、桐花树和无瓣海桑 5 种国内广泛分布的红树植物[28–29], 采用荟萃分析方法, 探究不同红树植物的最适盐度、最高耐受盐度以及最适淹水时间和最高耐受淹水时间。结合国内各宜林地和红树林修复规划区域的盐度和淹水条件, 筛选出适宜不同生境的造林树种, 有针对性地提出未来造林工程的应用对策, 以期可为红树林湿地恢复工程的树种选择以及幼苗保育提供科学依据。
1 材料与方法
1.1 数据来源
本研究检索了 Web of Science 数据库(1970—2020 年)和中国知网数据库, 英文检索主题词为“salinity”、“salt”、“NaCl”、“waterlogging”、“tide”、“sea level”、“inundate”+“Avicennia marina”、“Kan-delia obovata”、“Bruguiera gymnoihiza”, “Sonnera-tia apetala”和“Aegiceras corniculatum”。中文检索主题词为“盐度”、“淡水”、“NaCl”、“盐胁迫”、“耐盐”、“淹水”、“水位”、“水渍”、“潮汐”、“淹浸”+“白骨壤”、“秋茄”、“木榄”、“无瓣海桑”和“桐花树”。同时, 补充搜索了电子数据库检索到的文献中列出的参考文献。
1.2 选择标准
文献及数据纳入标准如下: 1)文献研究的红树植物种类包含要研究的目标红树植物; 2)文献中的实验包含探究盐度或淹水对红树植物影响, 或者盐度–淹水对红树植物的复合影响; 3)文献中包含以盐度、淹水时间或盐度–淹水时间为变量的实验部分, 其他条件在同一个实验中是相同的; 4)实验结果至少有一对数据。虽然不同的实验采用的红树植物的苗龄以及整个胁迫实验的时间不同, 但每个实验的其他条件以及施用的营养物质是合理的, 并不限制红树植物的生长。
1.3 数据提取与筛选
对于符合标准的文献, 提取其中的相关数据并建立电子数据库。主要提取信息包括: 实验材料信息、实验时间、盐度和淹水变量设定、实验结果数据以及论文名称、发表年月、作者和发表期刊。针对白骨壤、秋茄、桐花树、木榄和无瓣海桑这 5 种具有代表性的工程红树植物, 综合收集的数据从中选出 43 篇论文, 分别来自中国知网数据库(23 篇)和 Web of Science 数据库(20 篇)。为了充分地描述盐度和淹水胁迫对红树植物的影响, 选取苗高、叶片数、叶面积、基径、根长、总生物量、叶绿素含量、根活力、光合速率以及蒸腾速率等指标来表征红树植物的响应。对所有数据采用 Excel 2016软件进行统计, 用 Origin 2020 做箱型图来展示收集的数据。
1.4 统计学分析
使用 Stata 16.0 软件进行荟萃分析。使用 Q 检验(P 值)的定性分析和 I 2 检验的定量分析来判断纳入文献的异质性, 如果 P>0.1 且 I2<50%, 则表明没有异质性, 采用固定效应模型进行分析, 反之则采取随机效应模型进行分析。荟萃分析结果表明, 除淹水处理中的叶面积指标(P=0.000, I 2= 73.1%)外, 其他纳入分析的数据集均具有同质性(P>0.1 且 I 2< 50%), 说明相应研究中的盐度或淹水时间是影响红树植物生长的唯一因素, 其他实验条件不产生影响; 各指标对盐度或淹水时间的响应趋势相同(表 1)。
2 结果与分析
2.1 红树植物对盐胁迫的响应特征
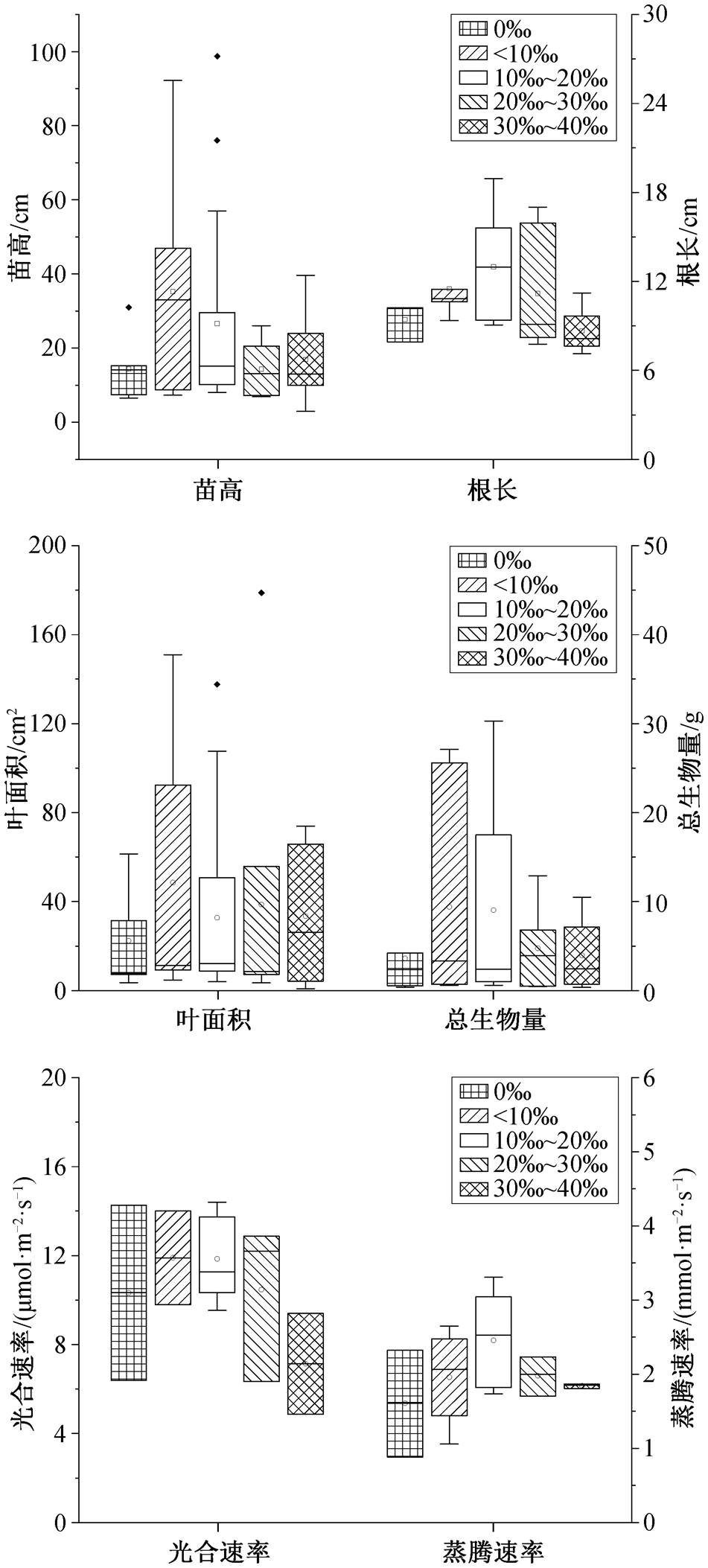
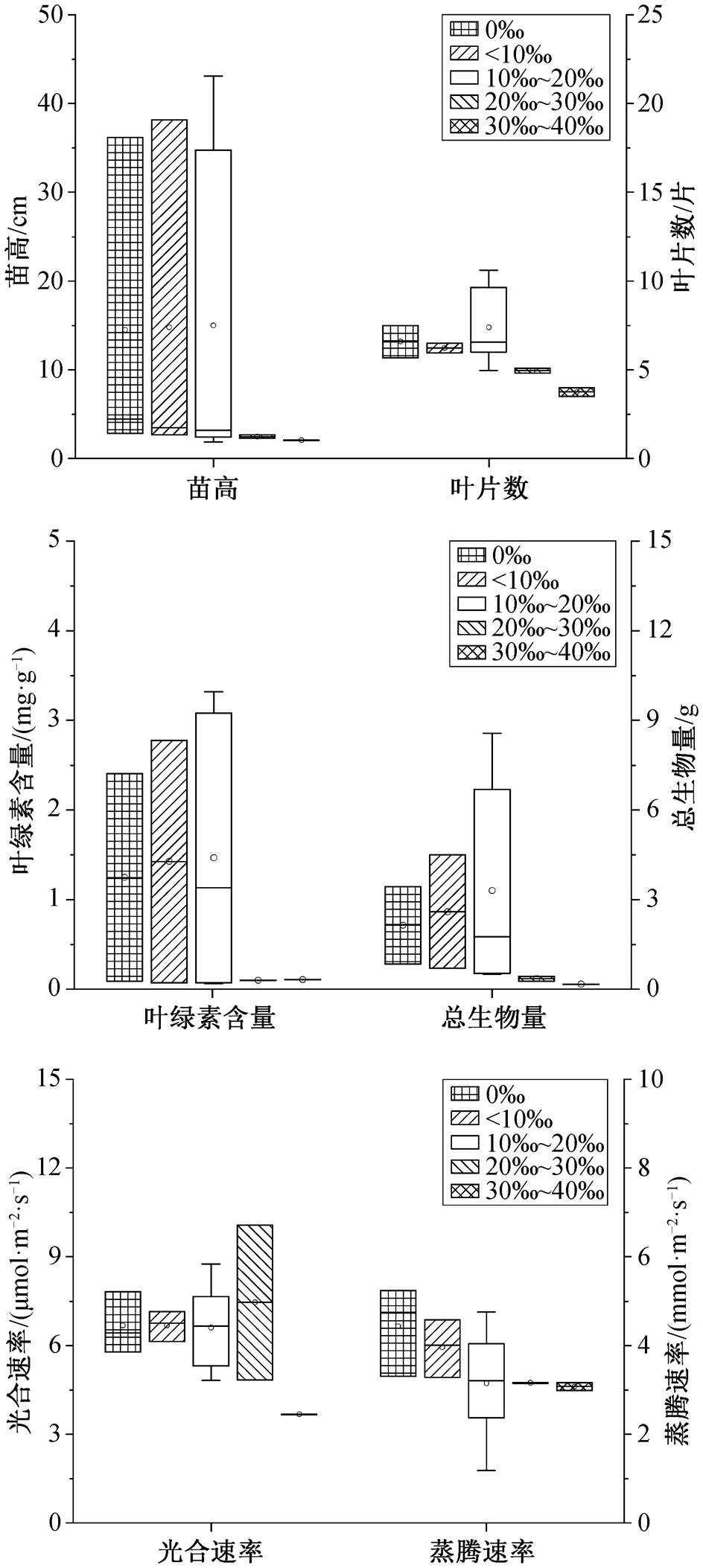
白骨壤的苗高、总生物量和光合速率在盐度小于 10‰的范围内达到最大值, 分别比对照组(淡水) 平均增加 132.6%, 42.2%和 15.2% (图 1)。在盐度10‰~20‰的范围内, 白骨壤的根长、叶面积和蒸腾速率达到最大值, 分别比对照组平均增加 27.5%, 51.7% 和 56.4%。在盐度 30‰~40‰的范围内, 与对照组相比, 其苗高、根长和光合速率分别平均降低8.0%, 20.0%和 31.0%, 较高的盐度会显著地抑制白骨壤生长, 并改变其根部结构[30]。因此, 白骨壤的最适盐度范围为 10‰~20‰, 最高耐受盐度范围为30‰~40‰, 其苗高、生物量、叶面积和光合速率对盐度的响应比较敏感。
表1 盐度和淹水对红树植物生理生化指标影响的统计检验结果
Table 1 Statistical test results of effects of salinity and water-logging on physiological and biochemical indices of mangrove plants
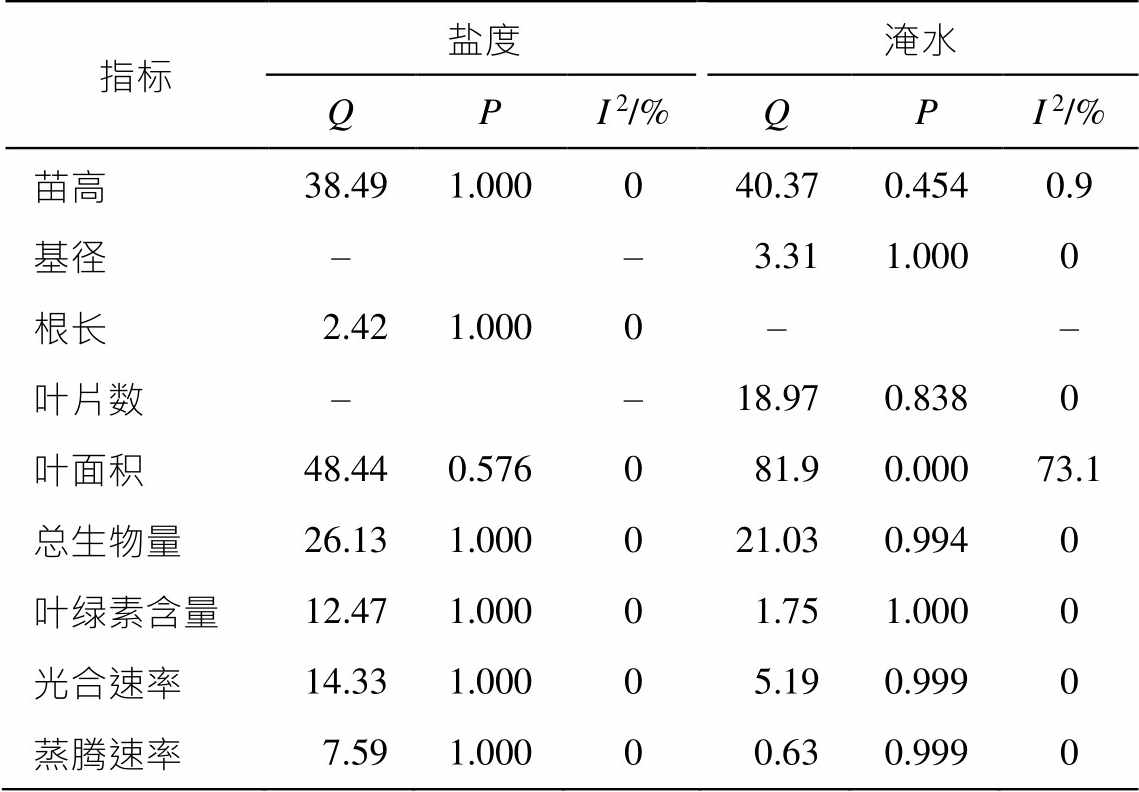
指标盐度淹水QPI2/%QPI2/% 苗高38.491.000040.370.4540.9 基径––3.311.0000 根长2.421.0000–– 叶片数––18.970.8380 叶面积48.440.576081.90.00073.1 总生物量26.131.000021.030.9940 叶绿素含量12.471.00001.751.0000 光合速率14.331.00005.190.9990 蒸腾速率7.591.00000.630.9990
秋茄的苗高、总生物量和光合速率在盐度小于10‰的范围内均达到最大值, 分别比对照组平均增加 28.2%, 508.1%和 51.8% (图 2)。在盐度 10‰~ 20‰的范围内, 叶绿素含量达到最大值, 比对照组平均增加 6.0%。在盐度 30‰~40‰的范围内, 与对照组相比, 秋茄的苗高和光合速率分别平均降低71.7%和 100%。因此, 秋茄的最适盐度范围在 10‰以内, 最高耐受盐度范围为 20‰~30‰, Ru 等[31]和邢建宏等[32]有相同的研究结果, 秋茄总生物量和光合速率对盐度的响应比较敏感。
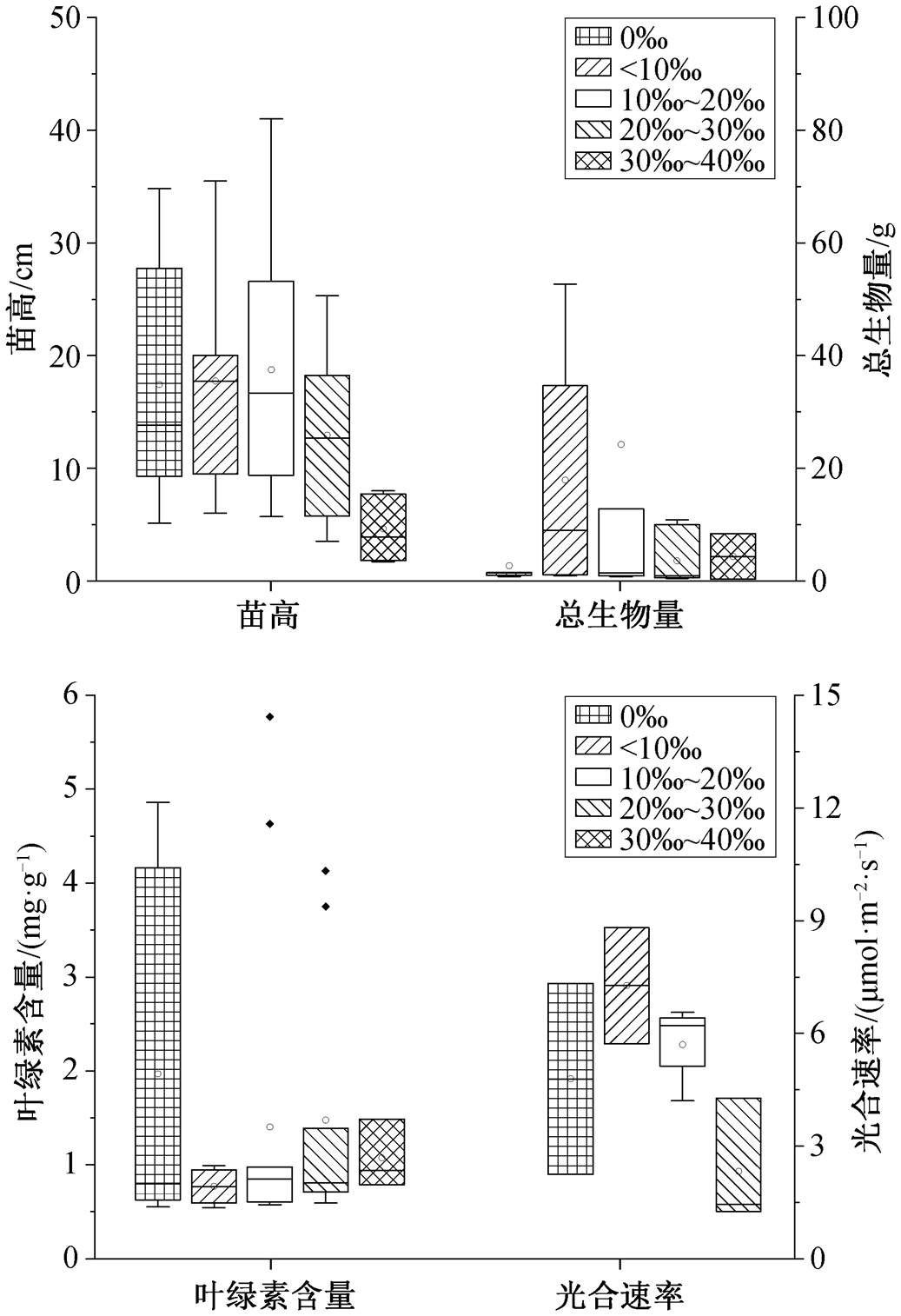
在盐度低于 10‰时, 桐花树的叶绿素含量最 高[33], 总生物量也达到最大值, 分别比对照组平均增加 67.1%和 11.1% (图 3)。在盐度 10‰~20‰的范围内, 苗高达到最大值, 比对照组增加 139.6%, 但总生物量和光合速率分别降低 30.3%和 26.6%。在盐度 20‰~30‰的范围内, 苗高和叶绿素含量分别降低 35.8%和 32.5%。因此, 桐花树的最适生长盐度在 10‰内, 最高耐受盐度范围为 20‰~30‰, Ke等[34]也有类似的研究结果, 其苗高和生物量对盐度的响应比较敏感。
在盐度小于 10‰的范围内, 木榄叶面积和总生物量达到最大值, 分别比对照组平均增加 123.1%和149.3% (图 4)。在盐度 10‰~20‰的范围内, 苗高达到最大值, 比对照组增加 26.7%, 叶面积和总生物量比对照组分别增加 36.9%和 70.5%。在盐度 20‰ ~30‰的范围内, 叶面积、叶绿素含量、光合速率和蒸腾速率与对照组相比分别降低 87.4%, 22.6%, 60.4%和 66.3%。木榄不是喜盐植物, 但具有一定的耐盐能力[35]。因此, 木榄最适盐度为10‰的范围以内, 最高耐受盐度为 10‰~20‰, 这与 Barik 等[36]和郑海雷等[37]的研究结果相同, 其叶面积、总生物量、光合速率和蒸腾速率对盐度的响应比较敏感。
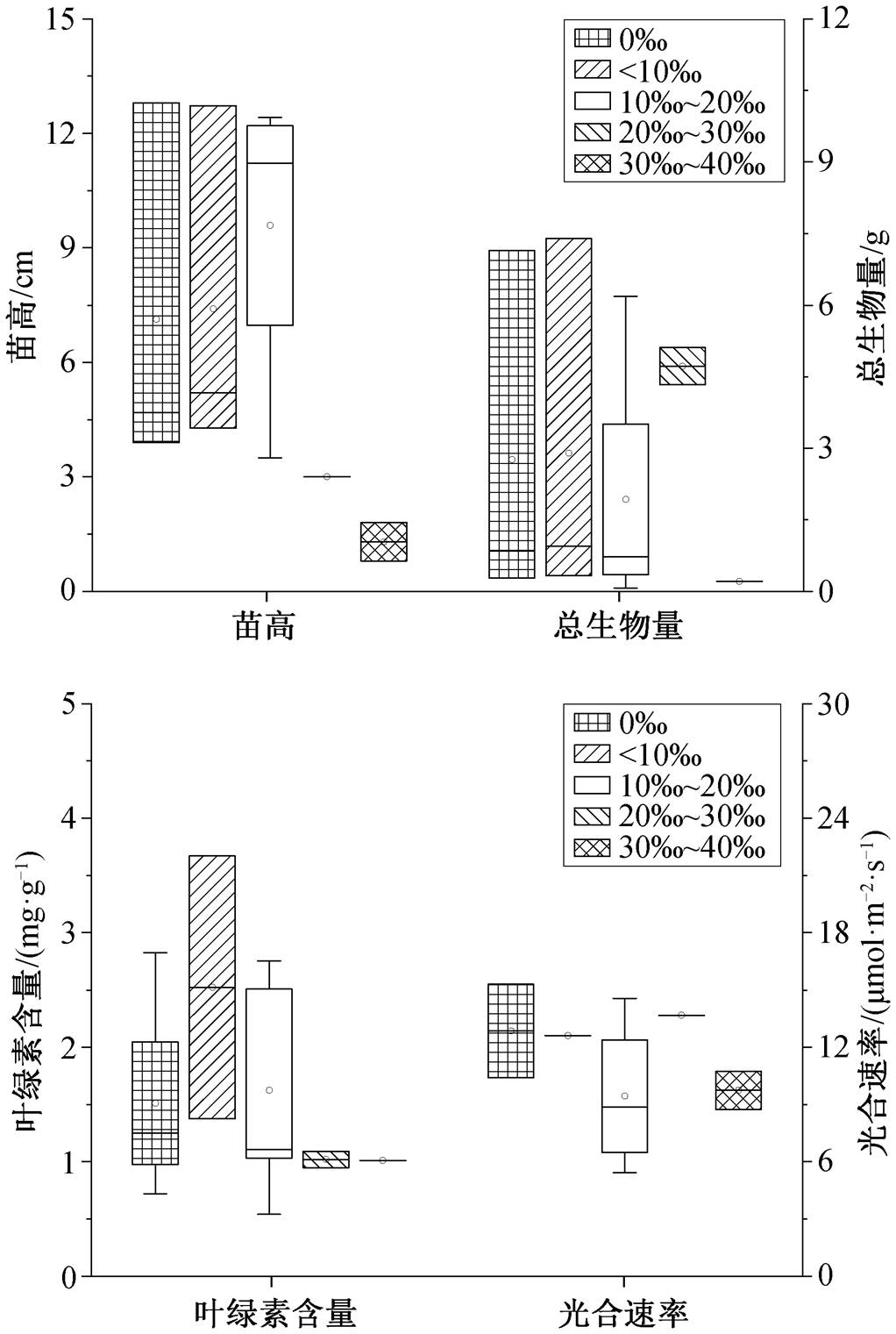
无瓣海桑的苗高、叶绿素含量和总生物量在盐度小于 10‰的范围内达到最大值, 分别比对照组平均增加 1.9%, 14.2%和 21.3% (图 5)。在盐度 10‰~ 20‰的范围以内, 叶片数达到最大值, 与唐密等[33] 的研究结果相同, 分别比对照组平均增加 12.1%。在盐度 20‰~30‰的范围内, 与对照组相比, 苗高、叶片数、叶绿素含量、总生物量和蒸腾速率平均降低83.0%, 24.8%, 92.0%, 83.3%和28.7%。因此, 无瓣海桑的最适盐度范围为 10‰以内, 最高耐受盐度为 10‰~20‰, Rahman 等[38]也有相同的研究结论, 苗高、叶绿素含量和总生物量对盐度的响应比较敏感。
综上, 所述 5 种红树植物对盐度的最高耐受值表现为白骨壤(40‰)>秋茄和桐花树(30‰)>木榄(20‰)>无瓣海桑(10‰); 白骨壤的最适盐度为10‰~20‰, 秋茄、桐花树、木榄和无瓣海桑的最适盐度为 10‰以内。盐度升高会导致植物体内 Na+和 K+等离子升高[39], 合成大量的有机物质(如氨基酸)作为调节渗透的保护剂[40–41], 提高其水分利用效率[42–43]。植物受到胁迫时会产生高浓度的 H2O2, 从而激活抗氧化防御系统[44]。例如, 随胁迫程度加剧, 桐花树根部中的过氧化物酶活性先上升后下降[45], 秋茄体内超氧化物歧化酶和过氧化氢酶活性则逐渐降低[46]。当环境盐分过高时, 植物的蒸腾速率将迅速下降[47], 并造成叶片早衰、光合面积减少以及叶绿体内羧化酶减少, 光合速率降低[47–48]; 同时, 丙二醛含量会显著上升, 破坏植物细胞膜, 造成植物新陈代谢紊乱[49]。
2.2 红树植物对淹水胁迫的响应特征
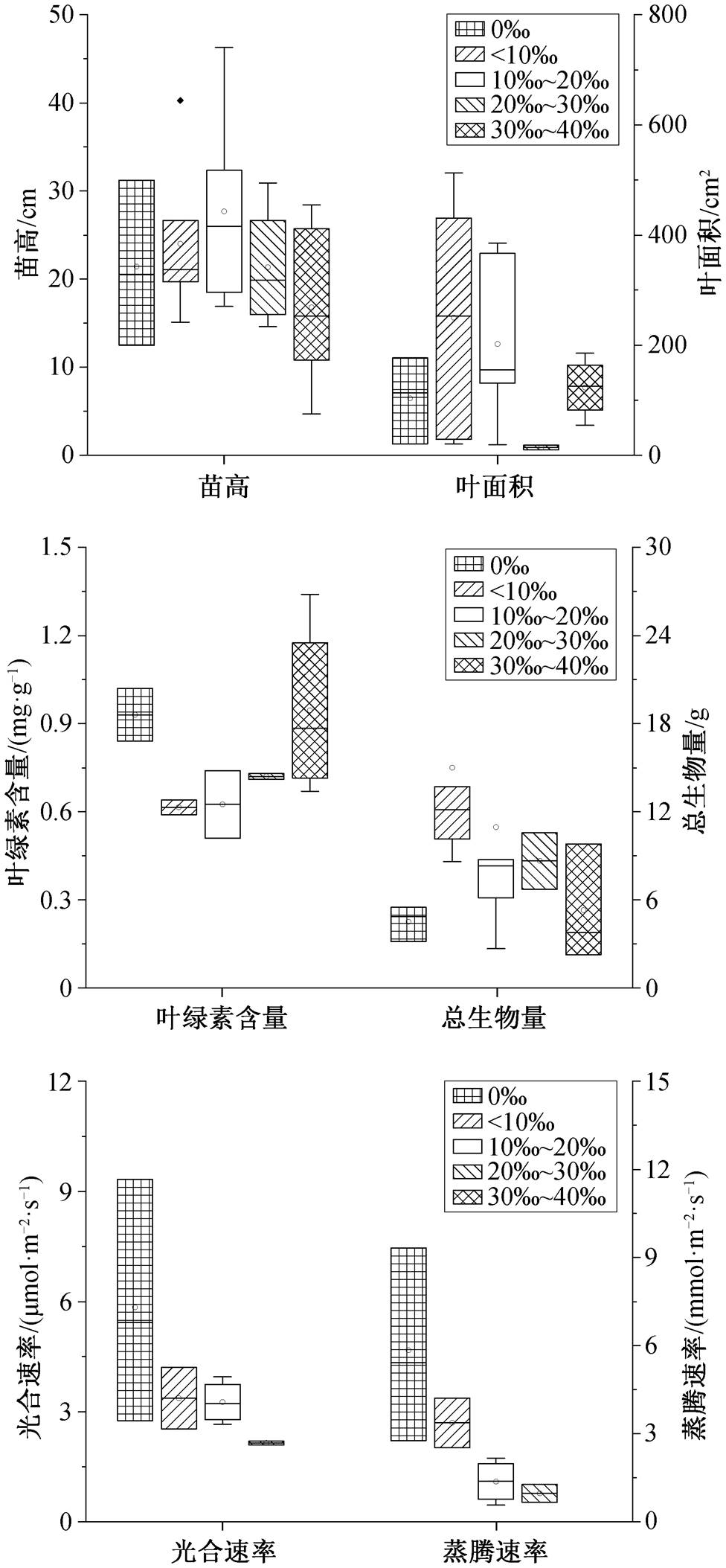
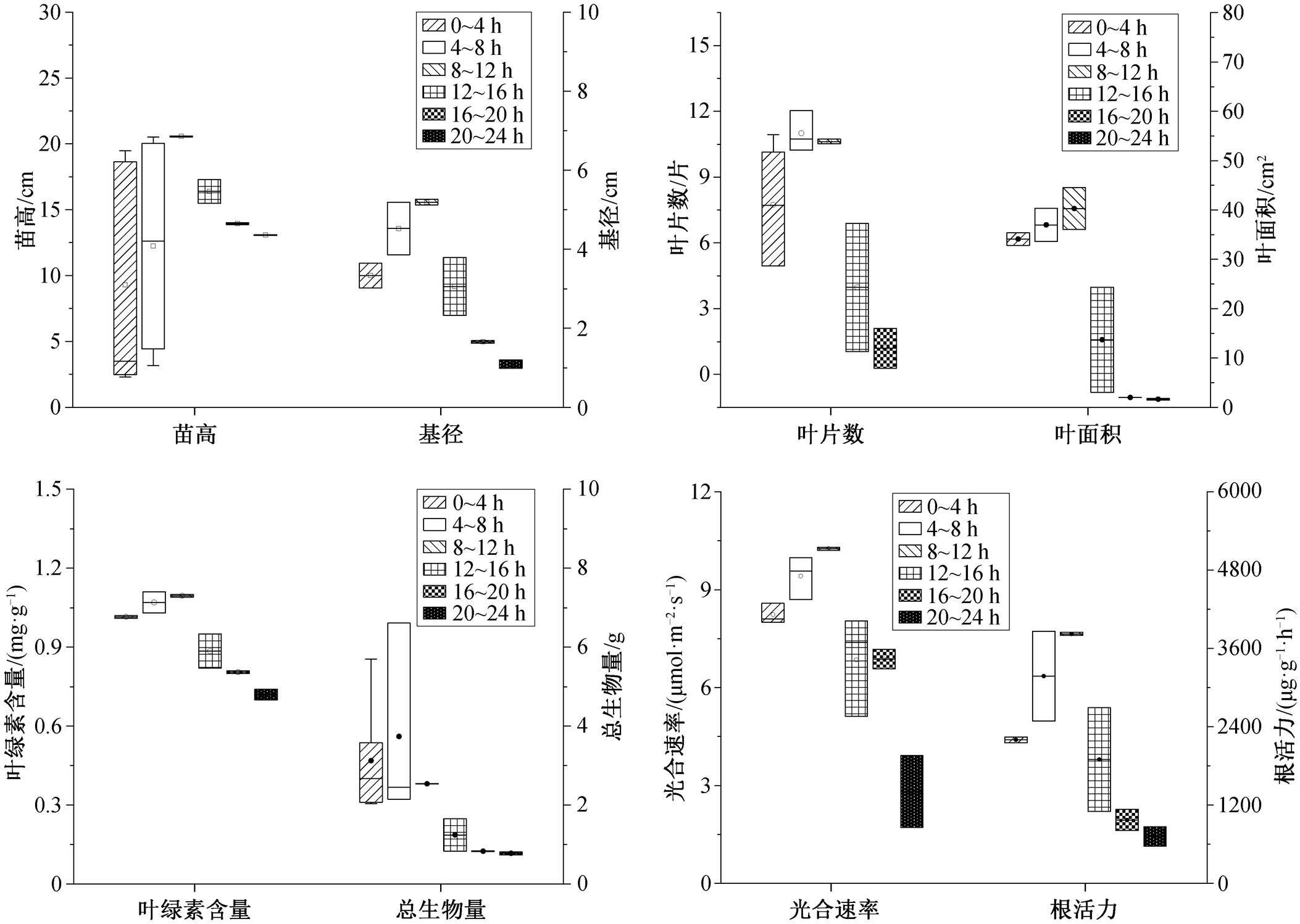
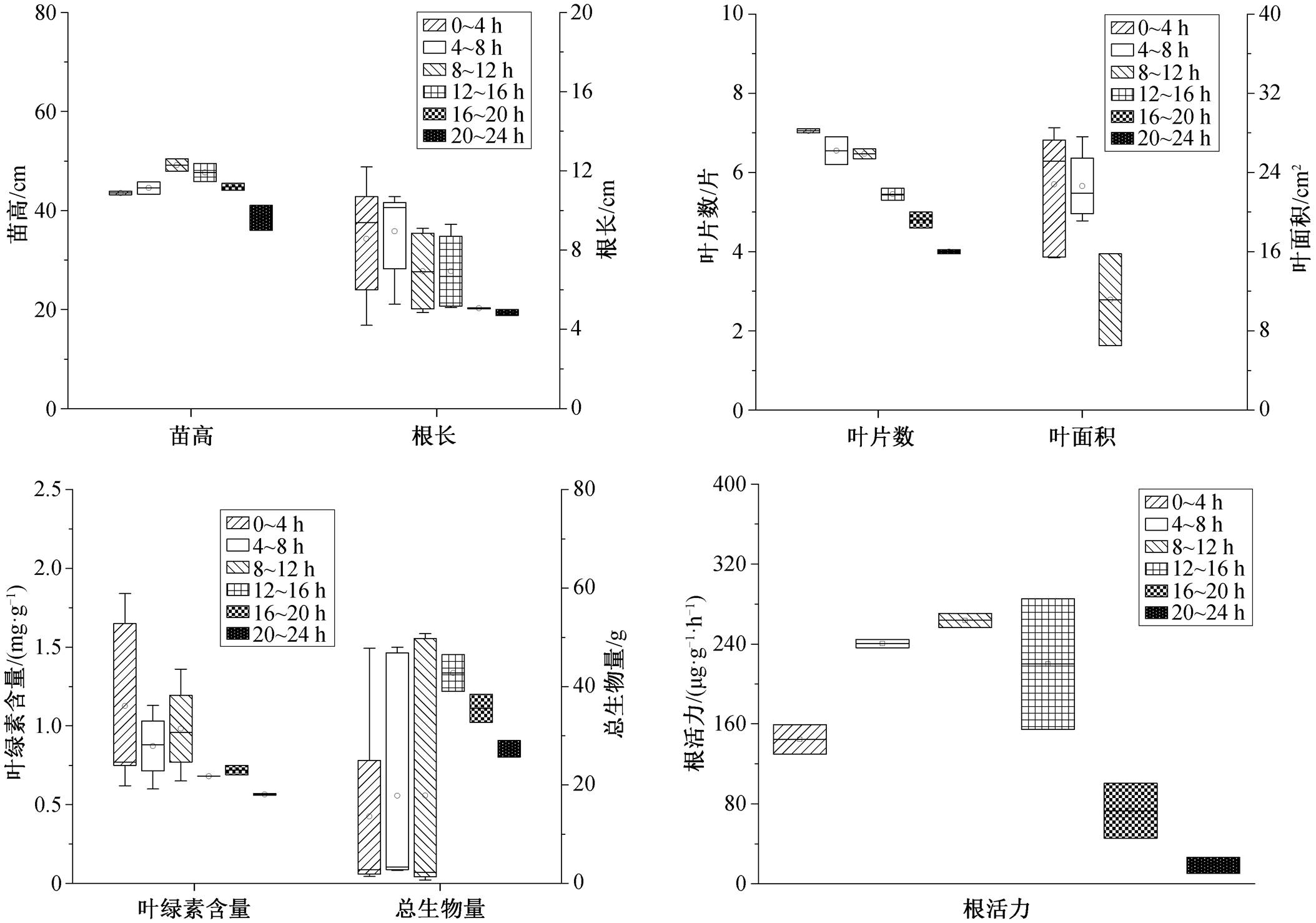
淹水时间为 4~8 小时时, 白骨壤叶片数达到最大值(图 6)。淹水时间为 8~12h 时, 苗高、基径、叶绿素含量、总生物量、光合速率和根活力达到最大值。淹水时间为 12~16h 时, 白骨壤的苗高、基径、叶片数、叶绿素含量、总生物量、光合速率以及根活力开始逐渐降低, 与最大值相比, 分别减低 20.3%, 45.4%, 3.4%, 19.2%, 51.1%, 33.0%和50.4%。淹水时间为 20~24 h 时, 各项指标分别下降 36.4%, 87.3%, 89.1%, 34.2%, 69.4%, 72.5%和81.2%。随淹水时间增加, 叶面积先增加后降低, 淹水时间为 8~12h 时达到最大值。白骨壤对淹水具有很强的耐受性[50], 因此每日最佳淹水时间是 8~12 h, 能够承受的最长淹水时间是 16 h。
淹水时间为 0~4h 时, 秋茄的叶片数达到最大值。淹水时间为 4~8h 时, 根长达到最大值(图 7)。淹水时间为 8~12h 时, 苗高、总生物量、叶绿素含量和根活力达到最大值。淹水时间为 12~16h 时, 叶片数、根长、叶绿素含量和根活力分别下降16.8%, 34.3%, 29.2%和 16.6%。当淹水时间为 20~ 24h 时, 各项指标分别下降 38.9%, 52.2%, 41.1%和 93.0%, 此时苗高下降 21.7%。叶面积随淹水时间的增加而降低, 淹水时间为 0~4h 时达到最大值。因此, 秋茄每日最佳淹水时间为 4~8h, 淹水时间在 8h 以内时生长良好, 刘滨尔等[51]有相同的研究结论, 秋茄能够承受的最长淹水时间是 12h, 超过 12 h 将不适合生长。
淹水时间为 8~12h 时, 桐花树的生物量、叶面积、叶片数、苗高和基径达到最大值[52], 此时, 其叶片的水分利用率也达到最大值[53]。12h 后, 各项指标开始下降。淹水超过 16 h 会影响桐花树根系硝酸还原酶活性, 降低根系中硝态氮和铵态氮含量, 影响其氮代谢[54]。在淹水条件下, 桐花树会提高根部孔隙度和气孔数量来保证氧气的运输和利用[55]。因此, 桐花树的每日最佳淹水时间为 8~12h, 能够承受的最大淹水时间为 16 h。
淹水时间在 12h 以内时, 木榄的基径逐渐增加, 在淹水时间在 16h 以内时, 其苗高逐渐增加[56]。淹水超过 12h后, 木榄的叶片结构会发生显著的变化, 导致光合作用降低以及水分运输减少[57]。因此, 木榄能够承受的每日淹水时间为 12h。
综上所述, 白骨壤和桐花树耐淹水能力较强, 每日最多能够承受 16h 的淹水, 其次是秋茄和木榄, 能够承受的最大淹水时间为 12h。
形态结构是反应植物对周围环境适应情况的重要指标[58], 水淹环境下根系活动受到限制[59], 此时红树植物倾向于产生轻质通气组织[60], 增加根系中的氮和磷为植物提供充足的养分[61]。短期淹水时, 红树植物体内水分运输和细胞结构保持相对稳定, 水分的增加可以提高叶片的叶绿素含量[62], 此时会产生抗氧化酶来抵抗淹水的伤害[63], 随着淹水时间的增加, 红树植物的水分储存、光合作用和叶肉电导率等功能将减弱[64], 合成大量丙二醛来损害细胞膜[65]。
2.3 红树植物对盐度–淹水复合胁迫的响应
不同物种的各项指标对盐度和淹水的敏感性存在差异[66]。淹水和盐胁迫都会降低植物的生产力, 在淹水条件下, 植物可能对盐度更加敏感[67]。若淹水或盐度处于红树植物的耐受范围内, 复合胁迫对红树植物不会造成显著的影响(表 2)。如当盐度为 7‰时, 秋茄的叶片数在淹水时间 4h 时达到最大值, 苗高在淹水时间 12h 时达到最大值[68], 与单一淹水胁迫的情况类似; 淹水时间为 8h 时, 秋茄的苗高、生物量以及光合速率在盐度为 5‰时达到最大值[69], 与单一盐分胁迫的情况类似。
表2 盐度‒淹水复合胁迫对红树植物的影响
Table 2 Effect of compound stress of salinity and waterlogging on mangroves
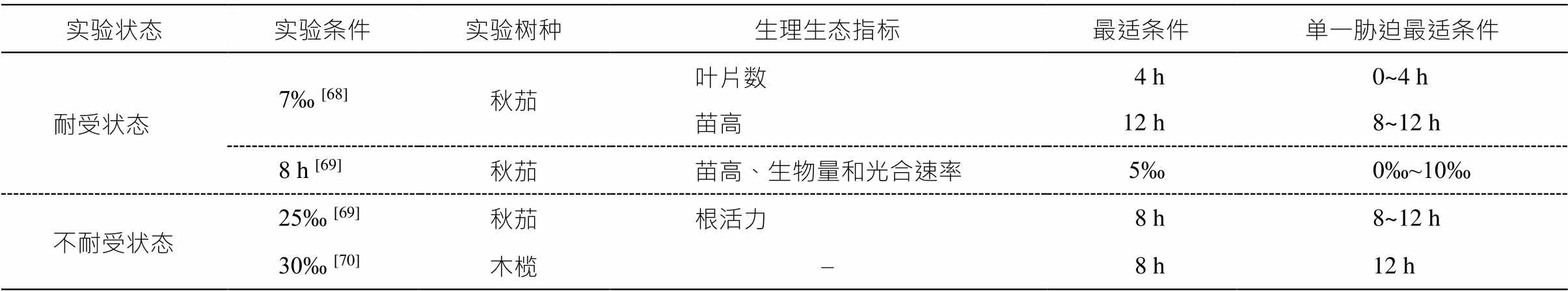
实验状态实验条件实验树种生理生态指标最适条件单一胁迫最适条件 耐受状态7‰ [68]秋茄叶片数4 h 0~4 h 苗高12 h8~12 h 8 h [69]秋茄苗高、生物量和光合速率5‰0‰~10‰ 不耐受状态25‰ [69]秋茄根活力8 h8~12 h 30‰ [70]木榄–8 h12 h
说明: 实验条件指盐度(‰)或淹水时间(h), “单一胁迫最适条件”是本文结论。
反之, 若超出其耐受范围时, 复合胁迫会导致红树植物的耐受性降低(表 2)。如当盐度为 25‰时, 秋茄根活力在每日淹水 8h 时达到最大值[69], 而仅淹水胁迫下为 14h[71]; 木榄在盐度为 30‰时, 能够承受 8h 的淹水[70], 而单一淹水胁迫时最高能够承受 12h 淹水。
2.4 应用对策
由于环境和气候的变化, 需要人工恢复红树 林[72]。目前, 从理论上讲, 中国红树林约有 16800hm2 的恢复潜力, 其中约 6400hm2 是用于水产养殖的池塘, 可作为红树林恢复的优先区域[73]。恢复潜力巨大的区域有海南省的三亚、青兰港和东寨港, 广西的北仑河口和铁山港[74], 广东省的阳江港、镇海湾、珠江口和红海湾等[75], 以及福建省的沙埕港、三沙湾、罗源湾和福清湾等处[76]。盐度的季节变化和平均值对红树植物形态和生理的影响程度相同[77]。因此, 在考虑盐度–淹水复合胁迫的情况下, 根据各省市最新发布的红树林修复保护规划, 结合各宜林地水文情况, 本文提出相应的造林物种建议(表 3)。值得注意的是, 选用无瓣海桑造林具有一定的争议性。碳储存是红树林造林的主要目标之一, 种植无瓣海桑可以迅速地增加红树林的面积和生物量[78], 但也可能阻碍本地物种的生长和发展, 改变本地植被的结构, 甚至影响本地红树林生态系统的功能, 因此在决定造林之前, 需要谨慎地考虑其入侵性[79–81]。
在红树林恢复过程中, 除需要将最常见的盐度和淹水因素纳入考虑外, 还需综合考虑其他环境影响因素, 如淡水流入及其周期、沉积物质地、有机质含量、温度、光照和风浪条件等。若种植区域寒害频发, 可选择耐寒的秋茄和桐花树[82]。若滩涂属于养分不充足的沙质沉积物, 可选择桐花树和白骨壤[83]。若需要在郁闭度较高的林下种植, 可选择耐荫的桐花树; 若种植区域的郁闭度不高, 则适合种植白骨壤、秋茄和木榄[84–85]。风浪较大的海岸, 适合种植高大速生的无瓣海桑; 其后缘风浪较小, 可种植本土红树植物[86]。
需要注意的是, 由于局部地形地貌的影响, 上述限制性环境因素会出现不同的组合, 红树林地理环境主要有以下几种类型[87]。
1)河流主导。这一类形受风浪影响较小, 河流带来的陆源沉积物沉积物营养充足, 影响因素主要是淡水流入及其周期。
2)潮汐主导。这一类型频繁受到潮汐淹浸, 影响因素主要是潮汐淹浸高度和淡水补充的缺乏。
3)波浪主导。这种类型的影响因素是沉积物以沙质为主, 同时受到高强度的海浪冲刷, 导致定植困难。
4)河流和波浪复合主导。这一类形包括海滩和泻湖, 影响因素是潮汐高度和海浪强度。
5)碳酸盐基质。这一类型的影响因素是土壤碳酸盐含量高, 钙化严重。因此, 需要因地制宜, 详细地考察以上因素在各宜林地的现实情况, 合理分配推荐树种的种植位置。
5 种红树植物的详细栽培方法见表 4。不同的树种适宜的种植方式不同, 大致分为两种。1)胚轴插植: 将红树植物胚轴直接或催芽后插植在造林地, 如秋茄和桐花树可采取这种方式。2)幼苗移植: 先将种子或胚轴在苗圃里育成小苗, 再将小苗移植到造林地, 如无瓣海桑可采取这种方式。造林地的位置选择也至关重要, 不同的树种适合生长在不同的潮位, 如桐花树和木榄适合种植在高、中潮位, 造林时要根据树种特性进行选择。为了提高人工林的生态稳定性和多样性, 需要根据不同树种的生态位和中间关系, 进行合理的物种搭配, 从而提高人工林的造林效率, 如秋茄、桐花树和白骨通壤常混交造林。
表3 红树林宜林地特征及造林树种推荐
Table 3 Characteristics of suitable afforestation land for mangrove and recommendation of mangrove species
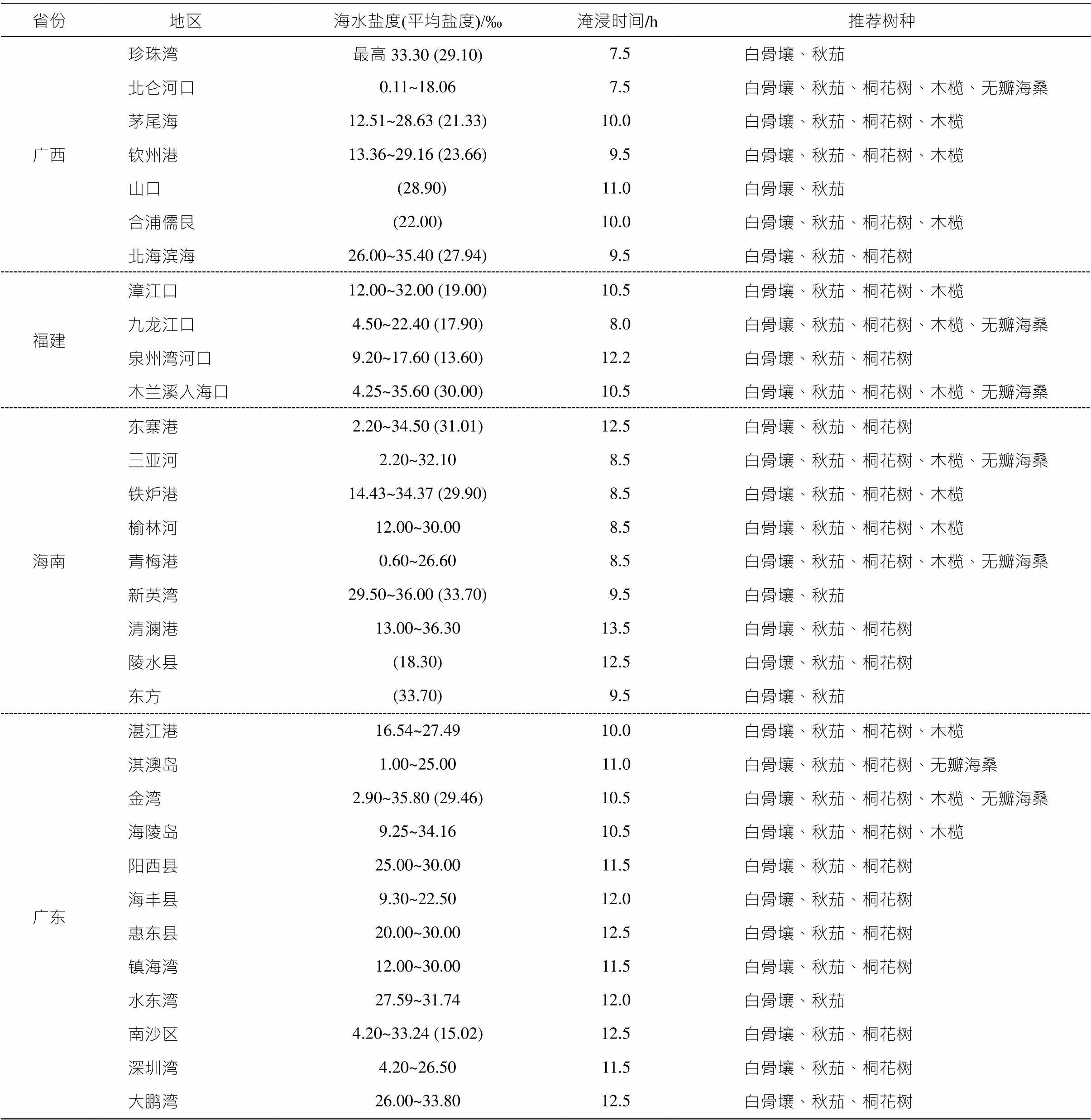
省份地区海水盐度(平均盐度)/‰淹浸时间/h推荐树种 广西珍珠湾最高33.30 (29.10)7.5白骨壤、秋茄 北仑河口0.11~18.067.5白骨壤、秋茄、桐花树、木榄、无瓣海桑 茅尾海12.51~28.63 (21.33)10.0白骨壤、秋茄、桐花树、木榄 钦州港13.36~29.16 (23.66)9.5白骨壤、秋茄、桐花树、木榄 山口 (28.90)11.0白骨壤、秋茄 合浦儒艮(22.00)10.0白骨壤、秋茄、桐花树、木榄 北海滨海26.00~35.40 (27.94)9.5白骨壤、秋茄、桐花树 福建漳江口12.00~32.00 (19.00)10.5白骨壤、秋茄、桐花树、木榄 九龙江口4.50~22.40 (17.90)8.0白骨壤、秋茄、桐花树、木榄、无瓣海桑 泉州湾河口9.20~17.60 (13.60)12.2白骨壤、秋茄、桐花树 木兰溪入海口4.25~35.60 (30.00)10.5白骨壤、秋茄、桐花树、木榄、无瓣海桑 海南东寨港2.20~34.50 (31.01)12.5白骨壤、秋茄、桐花树 三亚河2.20~32.108.5白骨壤、秋茄、桐花树、木榄、无瓣海桑 铁炉港14.43~34.37 (29.90)8.5白骨壤、秋茄、桐花树、木榄 榆林河12.00~30.008.5白骨壤、秋茄、桐花树、木榄 青梅港0.60~26.608.5白骨壤、秋茄、桐花树、木榄、无瓣海桑 新英湾29.50~36.00 (33.70)9.5白骨壤、秋茄 清澜港13.00~36.3013.5白骨壤、秋茄、桐花树 陵水县(18.30)12.5白骨壤、秋茄、桐花树 东方(33.70)9.5白骨壤、秋茄 广东湛江港16.54~27.4910.0白骨壤、秋茄、桐花树、木榄 淇澳岛1.00~25.0011.0白骨壤、秋茄、桐花树、无瓣海桑 金湾2.90~35.80 (29.46)10.5白骨壤、秋茄、桐花树、木榄、无瓣海桑 海陵岛9.25~34.1610.5白骨壤、秋茄、桐花树、木榄 阳西县25.00~30.0011.5白骨壤、秋茄、桐花树 海丰县9.30~22.5012.0白骨壤、秋茄、桐花树 惠东县20.00~30.0012.5白骨壤、秋茄、桐花树 镇海湾12.00~30.0011.5白骨壤、秋茄、桐花树 水东湾27.59~31.7412.0白骨壤、秋茄 南沙区4.20~33.24 (15.02)12.5白骨壤、秋茄、桐花树 深圳湾4.20~26.5011.5白骨壤、秋茄、桐花树 大鹏湾26.00~33.8012.5白骨壤、秋茄、桐花树
说明: 无瓣海桑人工造林可能具有生态风险[88], 请谨慎选择; 加粗地区因高盐度或长时间淹水需考虑盐度–淹水的复合胁迫效应。
表4 5种红树植物栽植方法
Table 4 Planting methods of five mangroves

造林树种栽植方法文献 白骨壤宜采取幼苗移植; 造林地选择低潮滩最宜; 宜与秋茄、桐花树、老鼠簕、红海榄等混交[29,89] 秋茄胚轴插植或幼苗移植均可; 造林地选择高、中、低潮滩均可, 高、中潮滩最适; 宜与桐花树、白骨壤等混交[90–91] 桐花树胚轴插植或幼苗移植均可; 造林地选择高、中潮滩最宜; 宜与白骨壤、秋茄、老鼠簕等混交[29,91–93] 木榄胚轴插植或幼苗移植均可; 造林地选择高、中潮滩最宜; 宜与海莲、角果木、红海榄、秋茄、桐花树等混交[29,91] 无瓣海桑宜采取幼苗移植; 造林地选择风浪小、盐度低的潮滩, 中、低潮滩最适; 宜与秋茄、木榄、红海榄等混交[91,94–95]
3 结论
1)5 种中国广泛分布的红树植物对盐度的最高耐受值如下: 白骨壤(40‰)>秋茄和桐花树(30‰)>木榄(20‰)>无瓣海桑(10‰), 白骨壤的最适盐度范围为 10‰~20‰, 秋茄、桐花树、木榄和无瓣海桑的最适盐度范围在10‰以内。
2)白骨壤和桐花树的耐淹水能力较强, 每日最多能够承受 16h 的淹水, 其次是秋茄和木榄, 能够承受的淹水时间为 12h。
3)当未超出红树植物的盐度或淹水耐受限度时, 盐度–淹水复合效应不显著。反之, 复合胁迫会降低红树植物的耐受性。
4)白骨壤和秋茄适应性较广, 在大多数宜林地都能生长, 桐花树和木榄次之。此外, 如需选无瓣海桑作为造林树种, 则应综合考虑树种优势及其生态风险。
参考文献
[1]Costa E S, Sa F, Gomes L E O, et al. Can severe drought periods increase metal concentrations in mangrove sediments? A case study in eastern Brazil. Science of the Total Environment, 2020, 748: 142443
[2]Senger D F, Hortua D A S, Engel S, et al. Impacts of wetland dieback on carbon dynamics: a comparison between intact and degraded mangroves. Science of the Total Environment, 2021, 753: 141817
[3]Donato D C, Kauffman J B, Murdiyarso D, et al. Mangroves among the most carbon-rich forests in the tropics. Nature Geoscience, 2011, 4(5): 293–297
[4]Du Jianguo, Hu Wenjia, Nagelkerken I, et al. Seagrass meadows provide multiple benefits to adjacent coral reefs through various microhabitat functions. Ecosys-tem Health and Sustainability, 2020, 6(1): 1812433
[5]Tomiczek T, Wargula A, Lomonaco P, et al. Physical model investigation of mid-scale mangrove effects on flow hydrodynamics and pressures and loads in the built environment. Coastal Engineering, 2020, 162: 103791
[6]Cahoon D R, Mckee K L, Morris J T. How plants influence resilience of salt marsh and mangrove wetlands to sea-level rise. Estuaries and Coasts, 2021, 44(4): 883–898
[7]Lee S Y, Primavera J H, Dahdou-Guebas F, et al. Ecological role and services of tropical mangrove ecosystems: a reassessment. Global Ecology and Bio-geography, 2014, 23(7): 726–743
[8]Goldberg L, Lagomasino D, Thomas N, et al. Global declines in human-driven mangrove loss. Global Change Biology, 2020, 26(10): 5844–5855
[9]Bryan-Brown D N, Connolly R M, Richards D R, et al. Global trends in mangrove forest fragmentation. Scientific Reports, 2020, 10(1): 7117
[10]Daza D A V, Moreno H S, Portz L, et al. Mangrove forests evolution and threats in the Caribbean Sea of Colombia. Water, 2020, 12(4): 1113
[11]Gilby B, Weinstein M P, Baker R, et al. Human actions alter tidal marsh seascapes and the provision of ecosystem services. Estuaries and Coasts, 2021, 44(6): 1628–1636
[12]Zhao Xiaochen, Rivera-Monroy V H, Wang Hongqing, et al. Modeling soil porewater salinity in mangrove forests (Everglades, Florida, USA) impacted by hydro-logical restoration and a warming climate. Ecological Modelling, 2020, 436: 109292
[13]卢元平, 徐卫华, 张志明, 等. 中国红树林生态系统保护空缺分析. 生态学报, 2019, 39(2): 684–691
[14]何斌源, 范航清, 王瑁, 等. 中国红树林湿地物种多样性及其形成. 生态学报, 2007, 27(11): 4859–4870
[15]廖宝文, 张乔民. 中国红树林的分布、面积和树种组成. 湿地科学, 2014, 12(4): 435–440
[16]Wang Wenqing, Fu Haifeng, Lee S Y, et al. Can strict protection stop the decline of mangrove ecosystems in China? From rapid destruction to rampant degrada-tion. Forests, 2020, 11(1): no. 55
[17]Jia Mingming, Wang Zongming, Zhang Yuanzhi, et al. Monitoring loss and recovery of mangrove forests during 42 years: the achievements of mangrove con-servation in China. International Journal of Applied Earth Observation and Geoinformation, 2018, 73: 535–545
[18]Syahid L N, Sakti A D, Virtriana R, et al. Determining optimal location for mangrove planting using remote sensing and climate model projection in southeast Asia. Remote Sensing, 2020, 12(22): 3734
[19]Wodehouse D C J, Rayment M B. Mangrove area and propagule number planting targets produce sub-optimal rehabilitation and afforestation outcomes. Estuarine Coastal and Shelf Science, 2019, 222: 91–102
[20]Devaney J L, Marone D, Mcelwain J C. Impact of soil salinity on mangrove restoration in a semiarid region: a case study from the Saloum Delta, Senegal. Resto-ration Ecology, 2021, 29(2): e13186
[21]Sharma S, Nadaoka K, Nakaoka M, et al. Growth performance and structure of a mangrove afforestation project on a former seagrass bed, Mindanao Island, Philippines. Hydrobiologia, 2017, 803(1): 359–371
[22]廖岩, 陈桂珠. 盐度对红树植物影响研究. 湿地科学, 2007, (3): 266–273
[23]Manea A, Geedicke I, Leishman M R. Elevated car-bon dioxide and reduced salinity enhance mangrove seedling establishment in an artificial saltmarsh community. Oecologia, 2020, 192(1): 273–280
[24]陈鹭真, 王文卿, 林鹏. 潮汐淹水时间对秋茄幼苗生长的影响. 海洋学报(中文版), 2005, 27(2): 141–147
[25]陈鹭真, 林鹏, 王文卿. 红树植物淹水胁迫响应研究进展. 生态学报, 2006, 26(2): 586–593
[26]Wang Wenqing, Li Xiaofeng, Wang Mao. Propagule dispersal determines mangrove zonation at intertidal and estuarine scales. Forests, 2019, 10(3): no. 245
[27]Xie Danghan, Schwarz C, Bruckner M Z M, et al. Mangrove diversity loss under sea-level rise triggered by biomorphodynamic feedbacks and anthropogenic pressures. Environmental Research Letters, 2020, 15 (11): 114033
[28]杨盛昌, 陆文勋, 邹祯, 等. 中国红树林湿地: 分布、种类组成及其保护. 亚热带植物科学, 2017, 46(4): 301–310
[29]郑俊鸣, 舒志君, 方笑, 等. 红树林造林修复技术探讨. 防护林科技, 2016(1): 99–103
[30]Zhao Yan, Wang Xun, Wang Youshao, et al. Effects of salt on root aeration, nitrification, and nitrogen uptake in mangroves. Forests, 2019, 10(12): 1131
[31]Ru Q M, Xiao Q, Lin P, et al. Short- and long-term effects of NaCl on physiological and biochemical characteristics in leaves of a true mangrove, Kandelia candel. Russian Journal of Plant Physiology, 2009, 56(3): 363–369
[32]邢建宏, 潘德灼, 谭芳林, 等. NaCl胁迫对秋茄幼苗根系抗氧化系统的影响. 热带亚热带植物学报, 2018, 26(3): 241–248
[33]唐密, 李昆, 向洪勇, 等. 盐胁迫对两种红树植物生态、生理及解剖结构的影响. 生态科学, 2014, 33(3): 513–519
[34]Ke Lin, Zhang Chunguang, Guo Chuling, et al. Effects of environmental stresses on the responses of mangrove plants to spent lubricating oil. Marine Pollution Bulletin, 2011, 63: 385–395
[35]Nandy Datta P, Dasgupta N, Das S. Differential expression of physiological and biochemical characters of some Indian mangroves towards salt tolerance. Physiology and Molecular Biology of Plants: An International Journal of Functional Plant Biology, 2009, 15(2): 151–160
[36]Barik J, Mukhopadhyay A, Ghosh T, et al. Mangrove species distribution and water salinity: an indicator species approach to Sundarban. Journal of Coastal Conservation, 2018, 22(2): 361–368
[37]郑海雷, 林鹏. 培养盐度对海莲和木榄幼苗膜保护系统的影响. 厦门大学学报(自然科学版), 1998, 37(2): 3–5
[38]Rahman M S, Sass-Klaassen U, Zuidema P A, et al. Salinity drives growth dynamics of the mangrove tree Sonneratia apetala Buch. -Ham. in the Sundarbans, Bangladesh. Dendrochronologia, 2020, 62: 125711
[39]靖姣姣, 张颖, 白志英, 等. 盐胁迫对小麦代换系渗透调节物质的影响及染色体效应. 植物遗传资源学报, 2015, 16(4): 743–750
[40]Dasgupta N, Nandy P, Sengupta C, et al. Salinity mediated biochemical changes towards differential adaptability of three mangroves from Indian Sundar-bans. Journal of Plant Biochemistry and Biotechno-logy, 2014, 23(1): 31–41
[41]Wang Lingxia, Pan Dezhou, Lv Xiaojie, et al. A multilevel investigation to discover why Kandeliacandel thrives in high salinity. Plant Cell and Environment, 2016, 39(11): 2486–2497
[42]陈坚, 李妮亚, 刘强, 等. NaCl处理下两种引进红树的光合及抗氧化防御能力. 植物生态学报, 2013, 37(5): 443–453
[43]Wang Wenqing, Xu Li, You Siyang, et al. Daily salinity fluctuation alleviates salt stress on seedlings of the mangrove Bruguiera gymnorhiza. Hydrological Processes, 2020, 34(11): 2466–2476
[44]Asaeda T, Barnuevo A. Oxidative stress as an indicator of niche-width preference of mangrove Rhizophora stylosa. Forest Ecology and Management, 2019, 432: 73–82
[45]许丽璇, 王春凤, 王慧云, 等. 桐花树抗重金属铅胁迫的谷胱甘肽及抗氧化酶作用. 中国农学通报, 2010, 26(15): 215–220
[46]王芳洲, 王友绍. Cu2+、Pb2+胁迫对秋茄幼苗可溶性蛋白和抗氧化酶活性的影响. 生态科学, 2020, 39 (4): 10–18
[47]Perri S, Entekhabi D, Molini A. Plant osmoregulation as an emergent water-saving adaptation. Water Re-sources Research, 2018, 54(4): 2781–2798
[48]He Yi, Yu Chenliang, Zhou Li, et al. Rubisco decrease is involved in chloroplast protrusion and Rubisco-containing body formation in soybean (Glycine max.) under salt stress. Plant Physiology and Biochemistry, 2014, 74: 118–124
[49]廖岩, 赵肖, 陈桂珠. 盐胁迫对无瓣海桑幼苗根茎叶膜保护系统的影响. 海洋环境科学, 2009, 28(2): 154–158
[50]Chen Yaping, Ye Yong. Early responses of Avicennia marina (Forsk.) Vierh. to intertidal elevation and light level. Aquatic Botany, 2014, 112: 33–40
[51]刘滨尔, 廖宝文, 方展强. 不同潮汐和盐度下红树植物幼苗秋茄的化学计量特征. 生态学报, 2012, 32(24): 7818–7827
[52]罗美娟, 张守攻, 崔丽娟, 等. 桐花树幼苗生长 与生物量分配对淹水胁迫的响应. 浙江林业科技, 2012, 32(4): 15–19
[53]罗美娟, 崔丽娟, 张守攻, 等. 淹水胁迫对桐花 树幼苗水分和矿质元素的影响. 福建林学院学报, 2012, 32(4): 336–340
[54]罗美娟. 淹水胁迫对桐花树幼苗氮代谢的影响. 福建林业, 2017(6): 29–32
[55]Cheng Hao, Wu Meilin, Li Changda, et al. Dynamics of radial oxygen loss in mangroves subjected to waterlogging. Ecotoxicology, 2020, 29(6): 684–690
[56]Xiao Yan, Wang Wenqing, Chen Luzhen. Stem ana-tomical variations in seedlings of the mangrove Bru-guiera gymnorrhiza grown under periodical water-logging. Flora, 2010, 205(8): 499–505
[57]Wang Wenqing, Xiao Yan, Chen Luzhen, et al. Leaf anatomical responses to periodical waterlogging in simulated semidiurnal tides in mangrove Bruguiera gymnorrhiza seedlings. Aquatic Botany, 2007, 86(3): 223–228
[58]黄依依, 蔡水花, 谭淑娟, 等. 真红树植物和半红树植物叶片性状的比较研究. 广西植物, 2020, 40 (3): 345–355
[59]宋香静, 李胜男, 郭嘉, 等. 环境变化对湿地植物根系的影响研究. 水生态学杂志, 2017, 38(2): 1–9
[60]Ye Y, Gu Y T, Gao H Y, et al. Combined effects of simulated tidal sea-level rise and salinity on seedlings of a mangrove species, Kandelia candel (L.) Druce. Hydrobiologia, 2010, 641(1): 287–300
[61]Ye Y, Tam N F Y, Wong Y S, et al. Does sea level rise influence propagule establishment, early growth and physiology of Kandelia candel and Bruguiera gym-norrhiza?. Journal of Experimental Marine Biology and Ecology, 2004, 306(2): 197–215
[62]沈小雪, 姜立得, 陶佳伟, 等. 退化秋茄林的补水修复实验研究. 北京大学学报(自然科学版), 2019, 55(4): 699–708
[63]Li Huan, Li Zan, Shen Zhijun, et al. Physiological and proteomic responses of mangrove plant Avicennia marina seedlings to simulated periodical inundation. Plant and Soil, 2020, 450: 231–254
[64]Xiao Yan, Jie Zuliang, Wang Mao, et al. Leaf and stem anatomical responses to periodical waterlogging in simulated tidal floods in mangrove Avicennia marina seedlings. Aquatic Botany, 2009, 91(3): 231–237
[65]伍卡兰, 彭逸生, 陈耿, 等. 非潮汐淹水对白骨壤膜脂过氧化系统的影响. 环境科学与管理, 2012, 37(8): 146–150
[66]Janousek C N, Dugger B D, Drucker B M, et al. Salinity and inundation effects on productivity of brackish tidal marsh plants in the San Francisco Bay-Delta Estuary. Hydrobiologia, 2020, 847(20): 4311–4323
[67]Janousek C N, Mayo C. Plant responses to increased inundation and salt exposure: interactive effects on tidal marsh productivity. Plant Ecology, 2013, 214(7): 917–928
[68]游惠明. 秋茄幼苗对盐度、淹水环境的生长适应. 应用生态学报, 2015, 26(3): 675–680
[69]廖宝文. 三种红树植物对潮水淹浸与水体盐度适应能力的研究[D]. 北京: 中国林业科学研究院, 2010
[70]黄丽, 谭芳林, 林捷, 等. 木榄幼苗对淹水‒盐胁迫的生理响应. 防护林科技, 2018(12): 1–4
[71]廖宝文, 邱凤英, 谭凤仪, 等. 红树植物秋茄幼苗对模拟潮汐淹浸时间的适应性研究. 华南农业大学学报, 2009, 30(3): 49–54
[72]Ellison A M, Felson A J, Friess D A. Mangrove rehabilitation and restoration as experimental adap-tive management. Frontiers in Marine Science, 2020, 7: no. 327
[73]Hu Wenjia, Wang Yuyu, Zhang Dian, et al. Mapping the potential of mangrove forest restoration based on species distribution models: a case study in China. Science of the Total Environment, 2020, 748: 142321
[74]Hu W, Wang Y, Dong P, et al. Predicting potential mangrove distributions at the global northern distri-bution margin using an ecological niche model: De-termining conservation and reforestation involvement. Forest Ecology and Management, 2020, 478: 118517
[75]晁碧霄, 胡文佳, 陈彬, 等. 基于MaxEnt模型的广东省红树林潜在适生区和保护空缺分析. 生态学杂志, 2020, 39(11): 3785–3794
[76]胡文佳, 晁碧霄, 王玉玉, 等. 基于最大熵模型 的福建省红树林潜在适生区评估. 中国环境科学, 2020, 40(9): 4029–4038
[77]Bompy F, Lequeue G, Imbert D, et al. Increasing fluctuations of soil salinity affect seedling growth performances and physiology in three Neotropical mangrove species. Plant and Soil, 2014, 380: 399–413
[78]Yu Chenxi, Feng Jianiang, Liu Kai, et al. Changes of ecosystem carbon stock following the plantation of exotic mangrove Sonneratia apetala in Qi’ao Island, China. Science of the Total Environment, 2020, 717: 137142
[79]Ren Hai, Guo Qinfeng, Liu Hong, et al. Patterns of alien plant invasion across coastal bay areas in southern China. Journal of Coastal Research, 2014, 30(3): 448–455
[80]Peng Yisheng, Diao Junming, Zheng Mingxuan, et al. Early growth adaptability of four mangrove species under the canopy of an introduced mangrove plan-tation: implications for restoration. Forest Ecology and Management, 2016, 373: 179–188
[81]Zhang Yasong, Yu Chenxi, Xie Junjie, et al. Comparison of fine root biomass and soil organic carbon stock between exotic and native mangrove. Catena, 2021, 204: 105423
[82]陈鹭真, 郑文教, 杨盛昌, 等. 红树林耐寒性和向海性生态系列对气候变化响应的研究进展. 厦门大学学报(自然科学版), 2017, 56(3): 305–313
[83]张乔民, 隋淑珍, 张叶春, 等. 红树林宜林海洋环境指标研究. 生态学报, 2001, 21(9): 1427–1437
[84]姜仲茂, 管伟, 丁功桃, 等. 不同光照和淹浸程度对木榄幼苗生长的综合效应. 生态环境学报, 2018, 27(10): 1883–1889
[85]谭淑娟, 李婷, 余素睿, 等. 光照强度对 8 种红 树植物幼苗生长和生物量分配的影响. 生态科学, 2020, 39(3): 139–146
[86]李玫, 廖宝文, 陈玉军, 等. 深水光滩红树林恢复工程技术——以珠海野狸岛为例. 林业科技通讯, 2021(2): 20–23
[87]Yuvaraj E, Dharanirajan K, Jayakumar S, et al. Geomorphic settings of mangrove ecosystem in South Andaman Island: a geospatial approach. Journal of Earth System Science, 2014, 123(8): 1819–1830
[88]Xie Wei, Zhong Cairong, Li Xinnian, et al. Hybridiza-tion with natives augments the threats of introduced species in Sonneratia mangroves. Aquatic Botany, 2020, 160: 103166
[89]陈伟, 钟才荣. 红树植物白骨壤的育苗技术. 热带林业, 2006, 34(4): 26–27
[90]黄良宙, 陈贰, 罗建华, 等. 湛江七种红树植物造林试验. 热带林业, 2019, 47(4): 44–47
[91]吴钟亲, 陈修仁, 田乐宇, 等. 退塘还林红树林混交造林技术. 林业科技通讯, 2021(2): 30–34
[92]廖宝文, 郑德璋, 郑松发, 等. 红树植物桐花树育苗造林技术的研究. 林业科学研究, 1998, 11(5): 23–29
[93]王杰瑶, 李金凤. 桐花树的育苗技术. 热带林业, 2009, 37(2): 30–31
[94]钟才荣, 李海生, 陈桂珠. 无瓣海桑的育苗技术. 广东林业科技, 2003, 19(3): 68–70
[95]李云, 郑德璋, 廖宝文, 等. 无瓣海桑引种育苗试验. 林业科技通讯, 1995(5): 21–22
Meta-analysis of Mangrove Salt-Waterlogging Tolerance and Application Strategies
JIANG Biqian, LI Ruili†, SHEN Xiaoxue, ZHANG Zhi, ZHANG Yueqi
School of Environment and Energy, Peking University Shenzhen Graduate School, Shenzhen 518055; † Corresponding author, E-mail: liruili@pkusz.edu.cn
Abstract The success of mangrove afforestation depends on the selection and plantation of mangrove species, in which salinity and waterlogging are important factors. In this study, based on literature retrieval and meta-analysis, we evaluated the salt and waterlogging tolerance of five mangrove species commonly used in domestic afforestation projects. The results showed that Avicennia marina was the most tolerant to salinity and waterlogging, which could grow in extreme salinity (40‰) and long-term waterlogging (16 h/d) site. Meanwhile, Kandelia obovata and Aegiceras corniculatum had the ability to withstand salinity and waterlogging, which could adapt to moderate salinity (30‰) and short-term inundation (12 h/d) environment. While Bruguiera gymnorrhiza and Sonneratia apetala were salt-sensitive, which grew better at low level of salinity (20‰) place, and the former could be planted in short-term immersion (12 h/d) area. When salinity or duration of immersion was out of the limit, compound stress would decrease the tolerance capabilities of mangroves. In the practical application of afforestation, A. marina, K. obovata and A. corniculatum were highly adaptable to aquatic environments with a high level of salinity and prolonged waterlogging, which were suitable afforestation species for mangrove wetland in southern China. This study also put forward the corresponding application strategies which contributes to the ecological restoration of mangroves in China.
Key words mangrove; salt tolerance; waterlogging tolerance; artificial afforestation; ecological restoration
doi: 10.13209/j.0479-8023.2022.062
深圳市自然科学基金重点(JCYJ20200109140605948)资助
收稿日期: 2021–08–23;
修回日期: 2021–09–22