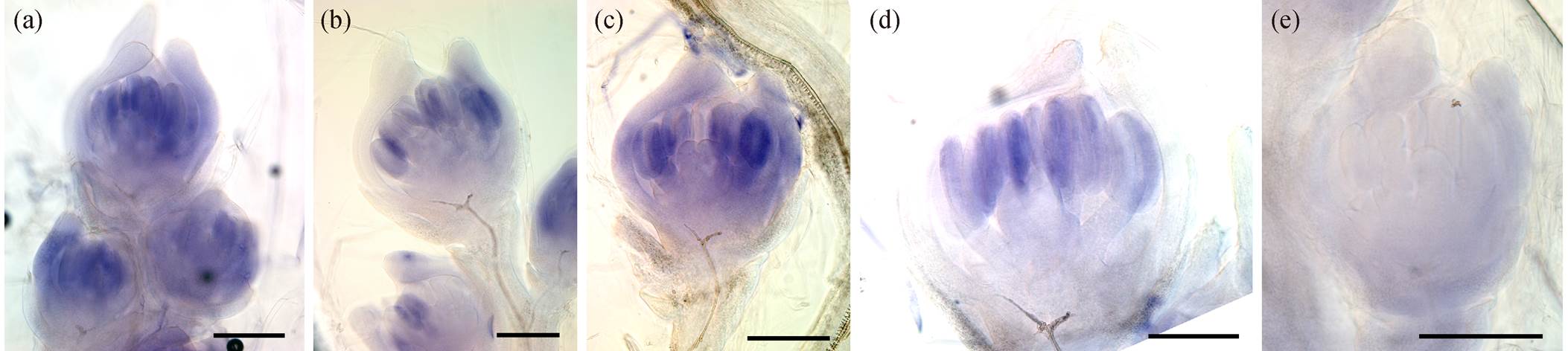
(a)~(d)为加水稻的SPW1/OsMADS16反义探针的杂交结果, (e)为加水稻的SPW1/OsMADS16正义探针后的杂交结果; 比例尺代表200 µm
图1 水稻SPW1/OsMADS16基因在小穗中的整体原位杂交结果
Fig. 1 Whole mount in situ hybridization results of SPW1/OsMADS16 gene in rice spikelets
摘要 以检测水稻 SUPERWOMAN1 (SPW1)/OsMADS16 基因的表达模式为例, 通过对杂交探针的制备、材料的固定及解离通透、显色等方面进行优化, 获得背景值低、特异性高的 SPW1/OsMADS16 基因特异性表达结果。建立一套针对水稻的整体原位杂交技术, 方法简单, 费用低廉, 可在离心管中多种材料同时进行。主要实验步骤包括探针制备、材料固定和解离、杂交反应、杂交后处理和检测显色等。提出的方法可为高通量检测水稻基因表达的时空模式及其功能分析提供技术条件。
关键词 整体原位杂交技术(WISH); 水稻; SUPERWOMAN1/OsMADS16基因
原位杂交(in situ hybridization, ISH)实验技术是分子生物学领域应用极为广泛的实验技术之一, 是研究植物发育过程中一种极为重要的技术手段。最初的原位杂交实验技术是 1969 年美国耶鲁大学Gall 等[1]用爪蟾核糖体基因氚标记的 RNA 探针与其卵母细胞的 DNA 杂交, 确定该基因定位于卵母细胞的核仁中, 是 DNA 的原位杂交方法[1–2]。根据探针的核酸性质不同, 可分为 DNA 探针、RNA 探针、cDNA 探针、cRNA 探针和寡核苷酸探针等。DNA 探针还有单链 DNA (single stranded, ssDNA)与双链 DNA (double stranded, dsDNA)之分, 检测目标也有 DNA 与 RNA 之分。原位杂交实验技术通过标记的探针与靶基因的专一性结合(核酸杂交配对), 精确定位组织内目标核酸, 从而展现在细胞, 甚至亚细胞水平下基因的时空表达情况。近几十年, 探针的标记方法由放射性向非放射性标记发展, 检出系统由化学染色向荧光发展, 检出目标由单一目标向多个即多色荧光发展, 灵敏度和分辨率也不断提高[3–4]。目前, 原位杂交实验技术仍然是基因表达分析的理想选择。
高度敏感的非放射性标记探针的积极发展, 使得原位杂交技术可直接在组织切片上进行, 但这种方式只能在二维平面上展现器官或组织中基因的表达情况, 对材料的大小也有很多限制。整体原位杂交实验技术(whole mount in situ hybridization, WISH) 是对完整的植物材料和组织进行探针杂交及检测,可以展现检测基因在组织器官中的三维分布。因此, WISH 成为了解基因在发育过程中时空表达状态, 特别是复杂基因表达模式(如某些只能在整个胚胎或整个组织器官层面进行分析的基因)的有力工具。该方法广泛应用于动物的胚胎和组织, 包括果蝇[3]、非洲蟾蜍[5]、鹌鹑[6]、粘液菌[7]和斑马鱼[8]等, 已成为一种胚胎学和发育生物学通用的方法。
与动物细胞相比, 植物细胞壁的存在使得探针穿透不足, 导致探针不易在植物组织中与靶基因配对。另外, 如果细胞或组织内检测的基因丰度较低, 并被相关的蛋白质掩盖保护, 都会增加检测的困难。成功的整体原位杂交实验技术需要在保持组织器官的形态完整与保证探针高效的渗透以便获得灵敏的检测结果之间进行平衡, 这就需要实验者的丰富经验。目前, 整体原位杂交实验技术在植物中成功地得到应用, 如拟南芥[9–10]、玉米[11]、黄瓜[12]和菊科植物欧蓍草[13], 但此项技术尚未在水稻中成功地使用。
在过去十多年中, 本实验室致力于构建和完善水稻早期雄蕊发育表达谱, 通过对水稻雄蕊早期发育过程的全基因组动态表达谱平台的构建, 完成对雄蕊早期发育过程的动态分子描述。通过对表达谱数据的挖掘, 发现共有 14571 个在雄蕊中表达的基因, 其中 2180 个基因在雄蕊早期发育过程中特异表达[14–15]。为了找到各个雄蕊发育时期特异基因的表达情况, 本研究优化整体原位杂交实验流程, 以便对水稻特异性表达雄蕊基因进行大规模的检测。本文以 SPW1/OsMADS16 为例, 介绍水稻整体原位杂交实验技术方法的改进情况。
实验中选用的材料是水稻(品种: ZH11)的幼穗, 材料由本实验室提供。地高辛检测和 NBT/BCIP 检测试剂盒购自美国罗氏公司, T7 和 Sp6 转录试剂盒购自美国 Promega 公司, 所有在杂交前和杂交中使用的器械、容器、移液管和枪头等均须无核酸酶(RNase 和 DNase)污染或经焦碳酸二乙酯(diethyl pyrocarbonate, DEPC)处理。
10×磷酸缓冲液(10×PBS): NaCl 80 g, KCl 2 g, Na2HPO4 14.4 g, KH2PO4 2.4 g, pH 7.4, 加ddH2O至1 L, 灭菌后使用。
4%多聚甲醛(paraformaldehyde, PFA)固定液: 用1×磷酸盐缓冲液(phosphate-buffered saline, PBS)配制, 4%多聚甲醛, 10%醋酸(acetic acid), 3%乙基苯基聚乙二醇(Nonidet P-40, NP-40), 15%二甲基亚砜(dimethyl sulfoxide, DMSO), 0.1%吐温-20 (Tween-20), 调节 pH至7.4, 60°C 放置溶解 2 小时后使用。
2×碳酸盐溶液(carbonate buffer): 将 80mM NaHCO3 和 120mMNa2CO3 溶解于焦碳酸二乙酯 (diethyl pyrocarbonate, DEPC)处理过的水中。
20×SSC溶液: NaCl 87.65 g, 柠檬酸钠 44.1 g, pH=7.0, 加水定容至500 mL, 灭菌后使用。
解离液: 50%去离子甲酰胺(formamide), 5×SSC, 3%十二烷基磺酸钠(sodium dodecyl sulfonate, SDS), 0.1%二硫苏糖醇(dithiothreitol, DTT), 0.1% Tween-20。
预杂交液: 50%去离子甲酰胺, 5×SSC, 3% SDS, 0.1% DTT, 0.1% Tween 20, 0.1 mg/mL Heparin, 1 mg/mL tRNA。
1.3.1探针制备
采用 RNA 探针, cRNA-RNA 之间形成的杂交体不受 RNA 酶影响。为提高探针的专一性, 用于转录 RNA 探针的 DNA 模板与靶序列以外的核酸序列无同源性。根据所检测基因的特异序列, 选取 200~ 400bp 片段设计相应引物, 在正向引物的 5'端加上SP6 启动子的核心序列(ATTTAGGTGACACTATA GAATA), 在反向引物的 5'端加上 T7 启动子的核心序列(AATTAATACGACTCACTATAGGG)合成引物。本实验所用的探针序列为: OM16insituT7: AAT TAATACGACTCACTATAGGGtgccgccctcaaggaggttcg 和 OM16insituSP6: ATTTAGGTGACACTATAGA ATAtaggccatgccgtgcaggttg。以纯化后的 cDNA 为模板, 在体外经 SP6 RNA 聚合酶或 T7RNA 聚合酶作用进行转录, 获得所需的探针: 变性后的模板 DNA 1µg, 地高辛标记物混合液 1µL, RNA 酶抑制剂 1µL, DTT 1µL, 5×转录缓冲液 2µL, T7 或 sp6 聚聚合酶 1µL, 加 ddH2O 至 10µL; 37°C 转录 2~4 小时。转录后的探针经紫外分光光度计测定浓度, 加入等体积的去离子甲酰胺分装存于−80°C 备用, 可以使用2~3 个月。
1.3.2材料的固定及去除叶绿素
使用 4%多聚甲醛固定液(4% paraformaldehyde fix solution, 4% PFA)固定水稻的小穗材料, 材料在真空抽气至无气泡产生为止。抽气完成后, 材料均沉在底部, 换新的 4%PFA 溶液, 4°C 固定过夜。4%PFA 是一种广泛用于固定组织、细胞等生物样品的溶液, 穿透力强, 固定均匀, 造成的组织损伤小, 较为温和, 可使细胞或组织的蛋白质凝固, 终止内源性或外源性酶反应, 防止组织自溶或异溶, 有原位保存抗原的作用, 避免抗原失活或弥散, 以便保持原有结构和形态。
对于含叶绿素的植物, 一般利用甲醇去除材料中的叶绿素。为了防止材料直接进入甲醇引起细胞形态的剧烈变化, 进行如下梯度置换: 磷酸盐缓冲液(PBST: 1×PBS溶液中加入 0.1% tween-20 溶液)、25%甲醇、50%甲醇、75%甲醇、100%甲醇、甲醇:乙醇(1:1)、纯乙醇、乙醇:二甲苯(1:1)、纯二甲苯、乙醇:二甲苯(1:1)和纯乙醇, 每步 30 分钟, 若材料体积较大, 渗透困难, 可延长处理时间。
1.3.3材料的解离及消化
要想获得高信噪比的杂交信号, 植物材料的解离是关键步骤。由于植物的细胞壁和细胞膜可以阻止探针侵入而与靶基因片段结合, 因此需要解离过程来增加材料的通透性而不破坏植物结构。用于增强组织的通透性和核酸探针的穿透性的试剂有很多, 如去污剂和蛋白酶K等。
将材料放于 55°C 解离液中, 处理 1~2 小时。这一步是为了让植物材料变得通透, 有利于探针的进入。然后用 4% PFA 室温固定 30 分钟, PBST 洗两次, 每次 20 分钟。用终浓度为 125μg/mL 的蛋白酶K 37°C 消化3 0 分钟, 4%的甘氨酸/PBST 处理 30 分钟以终止反应。10% DMSO/PBST 洗 2 次, 每次 10分钟。4% PFA 室温固定 30 分钟, PBST 洗 3 次, 每次 10 分钟。
1.3.4探针杂交
将材料加入适量预杂交液, 55°C 处理 2~4 小时, 如果材料较难渗透, 预杂交时间可延长。随后在预杂交液中加入 1μg/mL 左右的变性探针, 55°C 杂交24~48 小时。如果材料较难渗透, 探针杂交时间可延长。杂交后处理包括一系列不同浓度、不同温度盐溶液的漂洗。通过杂交后的洗涤可有效地降低背景信号, 获得较好的信噪比。用 2×SSCT 在 55°C 洗2 次, 每次 20 分钟, 去除未结合的探针。室温下PBST 洗 3 次, 每次 10 分钟。用 5% BSA/PBST 室温封闭 1 小时。按 1:2000 的比例, 在封闭液中加入DIG 抗体, 室温 16 小时或过夜温育。
杂交液中去离子甲酰胺的主要作用是调节杂交反应温度, 从而有助于保持组织的形态结构。去离子甲酰胺还可防止低温下非同源性片段的结合, 但具有破坏氢键的作用, 对核酸稳定性有影响。当杂交液含 50%甲酰胺且盐浓度在 0.75mol/L 左右时, RNA 探针的杂交温度是 50~55°C。
1.3.5显色观察
用 PBST 洗涤杂交后的水稻小穗, 然后用 NBT/ BCIP 溶液进行显色, 一般室温避光显色 12 小时, 在显色过程中注意观察, 达到要求后停止反应。反应结束后, 用梯度甲醇脱色, 洗去浮色。然后用三氯乙醛透明液处理 1 小时左右, 在显微镜下拍照。NBT/BCIP 是碱性磷酸酶最佳的底物组合之一, NBT 即四唑氮蓝(nitro-blue-tetrazolium), 为深蓝色无定形微溶物质; BCIP 即 5-溴-4-氯-3-吲哚-磷酸盐(5-bromo-4-chloro-3-indolyl-phosphate)。在碱性磷酸酶(AP)的催化下, BCIP 会被水解而产生强反应性产物。该产物与 NBT 发生反应, 形成不溶性的深蓝色至蓝紫色化学物。可根据颜色反应来确定目的蛋白的位置及表达情况, 从而显示靶基因的表达模式。
前人通过研究拟南芥的同源异型突变体, 提出植物花发育的 ABCDE 模型假说[16]: 在花中存在5 种类型的器官特异性基因功能区, 每个功能区分别控制相邻器官的发育, 决定雄蕊原基特征的是 B, C 和 E 类功能基因。其中, 水稻的 SPW1/OsMADS16属于花器官发育 ABCDE 模型中的 B 类基因, 起确定水稻花器官中浆片和雄蕊属性的作用, 与 APETAL-A3(AP3)/DEFICIENS (DEF)同源, 属于 MADS-box家族基因, MADS-box 基因在植物进化过程中对花结构的发育起到非常重要的作用[17–18]。SPW1/OsMA-DS16 与 DROOPING LEAF(DL)存在相互拮抗作用, 共同控制雄蕊和心皮的属性。SPW1/OsMADS16 与水稻 C 族基因 OsMADS3 和 OsMADS58 共同作用, 决定水稻花器官的形态[18], OsMADS16 基因功能缺失使得雄蕊转换成类似心皮结构的器官[19–21]。在此本文用 SPW1/OsMADS16 基因在水稻花序中的表达情况来验证整体原位杂交实验方法的可靠性。
本研究将正义探针作为阴性对照, 反义探针作为特异性的阳性探针进行整体原位杂交。如图 1 所示, 在水稻的小穗雄蕊中可见到清晰的杂交信号, 且背景基本上无染色。
为了验证整体原位杂交结果的准确性, 本研究同时进行 SPW1/OsMADS16 基因在水稻小穗的切片的原位杂交实验(图 2)。选取水稻小穗进行固定, 石蜡包埋后切片, 然后用与整体原位杂交一样的探针进行杂交。从图 2 可以看出, 切片原位杂交后NBT/BCIP 颜色在水稻雄蕊中显现, 与整体原位杂交实验的结果一致。SPW1/OsMADS16 基因在两种原位杂交实验中都表现出有水稻雄蕊特异性表达的模式, 与其是花器官发育的 B 类基因特征一致。
(a)~(d)为加水稻的SPW1/OsMADS16反义探针的杂交结果, (e)为加水稻的SPW1/OsMADS16正义探针后的杂交结果; 比例尺代表200 µm
图1 水稻SPW1/OsMADS16基因在小穗中的整体原位杂交结果
Fig. 1 Whole mount in situ hybridization results of SPW1/OsMADS16 gene in rice spikelets
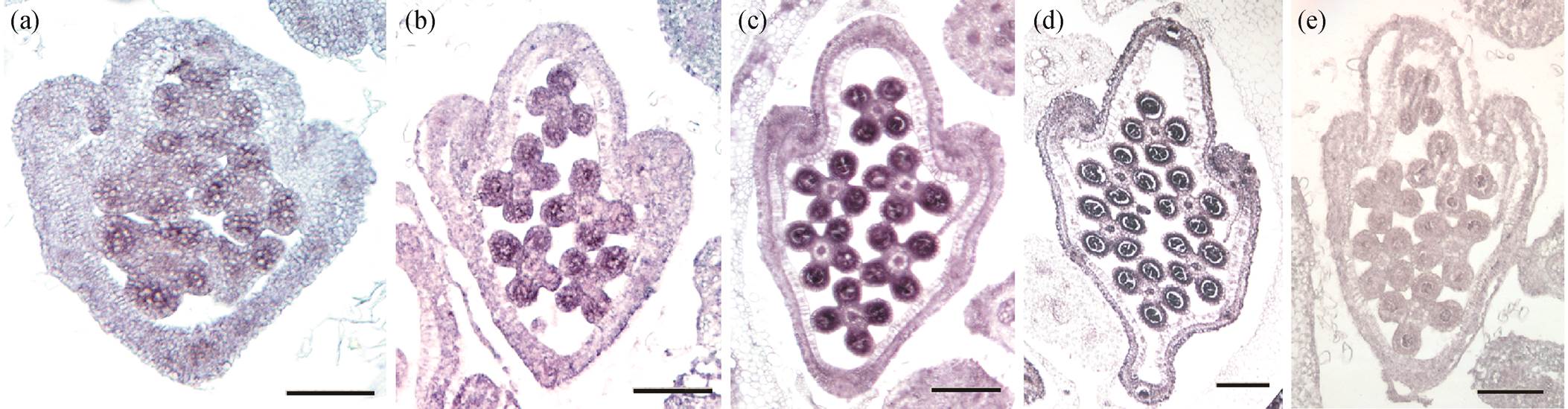
(a)~(d)为加水稻的SPW1/OsMADS16反义探针的杂交结果, (e)为加水稻的SPW1/OsMADS16正义探针后的杂交结果; 比例尺代表50 µm
图2 水稻SPW1/OsMADS16基因在小穗中的切片原位杂交结果
Fig. 2 In situ hybridization results of SPW1/OsMADS16 gene in rice spikelets
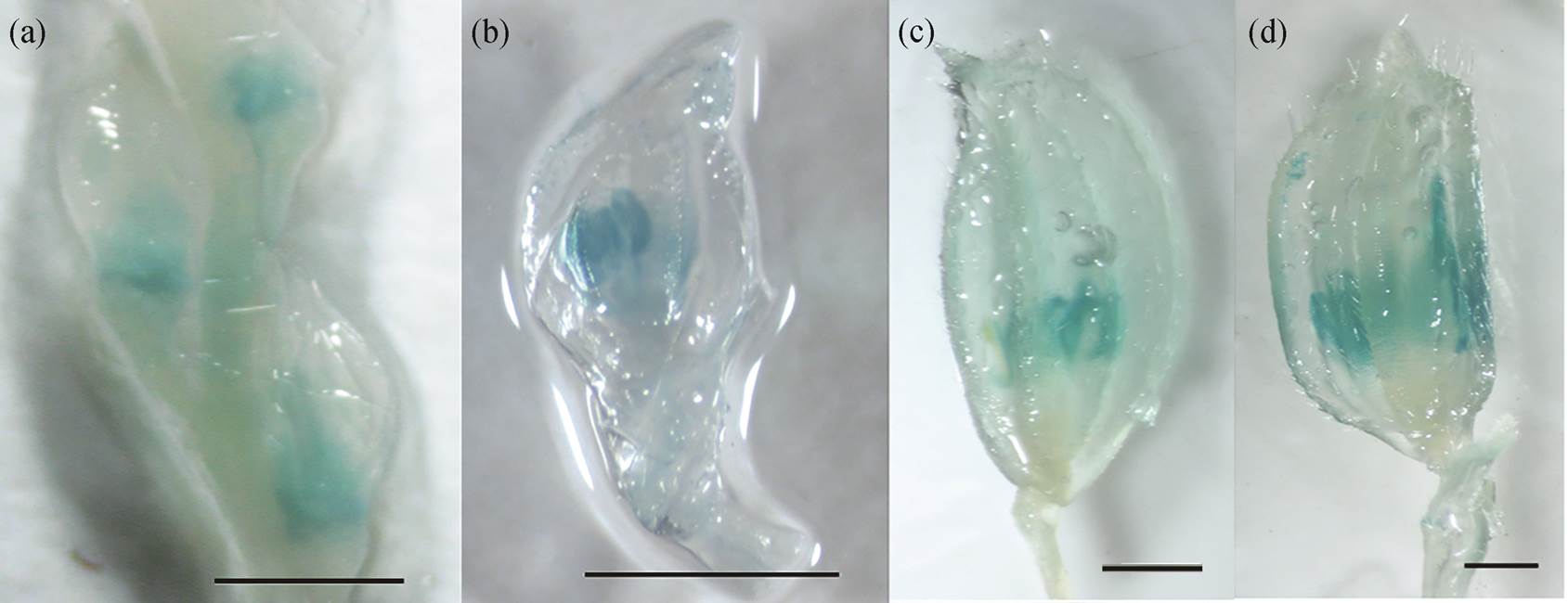
(a)~(d)为不同时期小穗GUS染色结果; 比例尺代表200 µm
图3 水稻SPW1/OsMADS16基因启动子驱动GUS在小穗中的染色结果
Fig. 3 Staining results of the GUS protein driven by the rice SPW1/OsMADS16 gene promoter in rice spikelets
同时, 本文设计 SPW1/OsMADS16 的启动子克隆引物 OM16 -SacI-F (CAAGAGCTCTGTCACTCAC CCCTGTAGTT)和 OM16 -KpnI-R (ATGGGTACCTC GATCTTGCCCCTCCCTAT), 以 SPW1/OsMADS16基因的上游 DNA 序列作为模板, 克隆了 SPW1/ OsMADS16 基因的上游 2057bp 作为启动子序列, 构建 SPW1/OsMADS16:GUS 载体, 进行水稻的转化实验。从 GUS 染色结果(图 3)可以看出, GUS 特异地在水稻小穗的雄蕊中显色。该结果不仅证明 SPW1/ OsMADS16 启动子在水稻雄蕊中的特异性, 同时验证了水稻整体原位杂交实验结果对于基因表达模式检测的准确性。
与动物细胞相比, 植物细胞中细胞壁的存在成为整体原位杂交实验过程中探针渗透的一个巨大的障碍。为增加通透性, 通常需要用蛋白酶 K 消化。如果消化程度不够, 会影响探针的渗透; 如消化过度则会破坏材料的结。因此, 消化的时间需要根据材料的幼嫩程度进行调整。为了增加组织通透性, 实验过程中可以使用 SDS, NP-40 和DMSO 等试剂来处理样品。同时, 为了保持器官或组织结构的完整性, 可以选择在消化后增加固定步骤。
另外, 有些植物的器官(如胚胎)中会积累大量黄酮或单宁物质, 用任何含甲醛的固定液来固定材料, 在组织中都会产生强烈的棕色背景。这是因为甲醛与许多植物酚, 特别是缩合单宁发生反应, 形成黑色聚合物, 这种黑色聚合物也会影响实验的结果。可选择使用荧光标记的探针取代化学染色的方法来解决这一问题。
探针的制备是整个原位杂交过程中一个重要的步骤, 制作良好探针的两个主要考虑因素是良好的模板序列和高效的标记反应。依据本实验室的经验, 原位杂交实验的理想模板序列长度应选取 300~ 1000bp, 然后用碳酸盐溶液(2×carbonate buffer)降解成小于 150 bp 的片段进行杂交效果会更好。一般的实验样品进行过夜杂交即可获得明显的信号, 但有时需要加长杂交时间(长达 3 天), 以便产生更强的信号, 此时需防止杂交过程中杂交液的蒸发, 以免产生背景干扰信号。
本研究从探针的制备、材料解离通透时间的控制和底物显色等环节进行优化, 简化操作步骤, 降低实验成本, 提高显色特异性, 从而建立一种高效、实用的水稻整体原位杂交技术, 为水稻基因表达的时空模式检测以及功能分析提供了技术条件。
与基于切片的组织原位杂交实验技术相比, 整体原位杂交不仅更适用于不便进行切片的植物材料(如幼嫩的组织和器官), 而且实验流程相对简单, 试剂用量少, 可以在短时间内进行高通量的基因检测工作。结合自动化仪器, 整体原位杂交实验技术会是一种可以高通量地检测基因时空表达模式的有效方法。
参考文献
[1] Gall J G, Pardue H L. Formation and detection of RNA-DNA hyvride molecules in the xytological pre-paration. PNAS, 1969, 63(2): 378–381
[2] John H A, Birstiel M L, Jones K W. RNA-DNA bybrids at the cytological level. Nature, 1969, 223: 582–587
[3] Tautz D, Pheifle C. A non-radioactive in situ hybri-dization method for the localization of specific RNAs in Drosophila embryos reveals translational control of the segmentation gene hunchback. Chromosoma, 1989, 98(2): 81–85
[4] Hougaard D M, Hansen H, Larsson L I. Non-radioac-tive in situ hybridization for mRNA with emphasis on the use of oligodeoxynucleotide probes. Histoche-mistry and Cell Biology, 1997, 108(4): 335–344
[5] Hemmati-Brivanlou A, Frank D, Bolce M E, et al. Localization of specific mRNAs in Xenopus embryos by whole-mount in situ hybridization. Development, 1991, 110(2): 325–330
[6] Coutinho L L, Morris J, Ivarie R. Whole mount in situ detection of low abundance transcripts of the myo-genic factor qmf1 and myosin heavy chain protein in quail embryos. BioTechniques, 1992, 13(5): 722–724
[7] Escalante R, Loomis W F. Whole-mount in situ hybri-dization of cell-type-specific mRNAs in Dictyoste-lium. Developmental Biology, 1995, 171(1): 262–266
[8] He J B, Mo D S, Chen J Y, et al. Combined whole-mount fluorescence in situ hybridization and antibody staining in zebrafish embryos and larvae. Nature Protocols, 2020, 15(10): 3361–3385
[9] GarcÍA-Aguilar M, Dorantes-Acosta A, PÉRez-Es-paña V, et al. Whole-mount in situ mRNA localization in developing ovules and seeds of Arabidopsis. Plant Molecular Biology Reporter, 2005, 23(3): 279–289
[10] Hennig L, Köhler C. Plant Developmental biology. Totowa, NJ: Humana Press, 2010
[11] Cordts S, Bantin J, Wittich P E, et al. ZmES genes encode peptides with structural homology to defen-sins and are specifically expressed in the female gametophyte of maize. Plant Journal, 2001, 25(1): 103–114
[12] Sun J J, Li F, Wang D H, et al. CsAP3: a cucumber homolog to Arabidopsis APETALA3 with novel cha-racteristics. Frontier in Plant Science, 2016, 7: 1181
[13] Sha S, Chen D, Liu M, et al. To be serrate or pinnate: diverse leaf forms of yarrows (Achillea) are linked to differential expression patterns of NAM genes. Annals of Botany, 2017, 121(2): 255–266
[14] Lu X C, Gong H Q, Huang M L, et al. Molecular analysis of early rice stamen development using organ-specific gene expression profiling. Plant Mole-cular Biology, 2006, 61(6): 845–861
[15] Chen R, Shen L P, Wang D H, et al. A gene expres-sion profiling of early rice stamen development that reveals inhibition of photosynthetic genes by OsMADS58. Molecular plant, 2015, 8(7): 1069–1089
[16] Theissen G , Saedler H. Floral quartets. Nature, 2001, 409: 469–471
[17] Nagasawa N, Miyoshi M, Sano Y, et al. SUPERW-OMAN1 and DROOPING LEAF genes control floral organ identity in rice. Development, 2003, 130(4): 705–718
[18] Yun D P, Liang W Q, Drenic L, et al. OsMADS16 genetically interacts with OsMADS3 and OsMADS58 in specifying floral patterning in rice. Molecular Plant, 2013, 6(3): 743–756
[19] Xiao H, Wang Y, Liu D F, et al. Functional analysis of the rice AP3 homologue OsMADS16 by RNA inter-ference. Plant Molecular Biology, 2003, 52(5): 957–966
[20] Lee S, Jeon J, An K, et al. Alteration of floral organ identity in rice through ectopic expression of OsMADS16. Planta, 2003, 217(6): 904–911
[21] Yoshida H, Nagato Y. Flower development in rice. Journal of Experimental Botany, 2011, 62(14): 4719–4730
Improvement of Experimental Techniques of Whole Mount in situ Hybridization Technology in Rice
Abstract Taking rice SUPERWOMAN1 (SPW1)/OsMADS16 gene as an example, by optimizing the preparation of hybridization probes, material immobilization, dissociation, permeabilization and color development, the SPW1/OsMADS16 gene expression patterns with high specificity but low backgrounds were obtained. The procedure of whole mount in situ hybridization (WISH) technology for rice is simple and inexpensive, and it can be performed simultaneously with several species in the centrifuge tube. The main experimental steps include probe preparation, material immobilization and dissociation, hybridization reaction, post-hybridization treatment and detection and color development. The proposed method provides a technological basement for a protentional high throughput detection of spatiotemporal gene expression patterns and functional analysis in rice.
Key words whole mount in situ hybridization (WISH); rice; SUPERWOMAN1/OsMADS16
doi: 10.13209/j.0479-8023.2022.019
蛋白质与植物基因研究国家重点实验室开放运行费(8102000374/006)资助
收稿日期: 2021-07-06;
修回日期: 2021-09-15