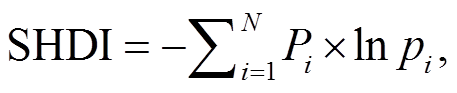 (1)
(1)摘要 以不透水面占比作为城市化强度(UI)的定量指标, 采用全球哺乳动物丰富度网格数据集, 结合气候因素和生境特点, 研究我国长三角城市群哺乳动物丰富度沿城乡梯度的空间分布格局。通过对比长三角城市群及其所处自然地理区(华东)哺乳动物丰富度的分布格局和驱动因素, 分析城市化和背景气候的贡献。结果表明: 1) 哺乳动物丰富度沿城乡梯度均呈水平“S”型分布; 2)在背景自然地理区域, 气候因素(如年平均降水和最冷月最低温等)对丰富度分布起主导作用(R2=88.2%); 3)UI 在华东地区与丰富度负相关, 在长三角城市群则弱正相关(P<0.001), 林地对丰富度存在正面影响, 耕地存在负面影响。因此, 城市化对哺乳动物丰富度影响在城市群尺度更为明显, 应更多关注如何区分城市化和背景气候对生物多样性的影响。
关键词 城乡梯度; 哺乳动物; 生物多样性; 不透水表面; 城市化强度; 背景气候
生物多样性是人类社会赖以生存发展的重要物质基础[1], 其间接价值往往与保持水土、调节气候及维持自然平衡等重要生态系统服务紧密相连。根据世界自然保护联盟的统计, 目前全球有超过28000 种物种濒临灭绝, 其中哺乳类占 1/4。城市化是对地表改变最强烈且不可逆转的人类活动, 被广泛认为是造成全球生物多样性丧失和生物同质化的重要原因之一[2]。全球超过一半的人口居住在城市, 预计 2030 年, 全球的城市化土地面积将增加一倍, 并且未来的城市扩张将主要发生在全球生物多样性热点区(如中国)。对于小型的自然保护区而言, 其生物多样性更容易受到城市扩张的威胁[3–4]。随着全球城市化侵占越来越多的自然栖息地, 城市已然变成野生动物的另一重要栖息地, 近 20 年来, 学者们对城市野生动物多样性的关注也日益加强, 但目前的研究集中在北美和欧洲等发达国家和地区, 只有极少数研究关注南美、非洲和亚洲地区, 这些地区不仅包含众多的生物多样性热点, 同时正在经历快速的城市化进程[5]。未来全球变化(如气候变暖)将如何影响自然生态系统中各种生态和进化过程, 一直是生态学家关注的焦点问题, 而城市正是可用于替代研究大空间、长时间尺度问题的绝佳实验场所[6], 研究城市化对生物多样性的影响将有助于窥见并应对未来全球变化对全球生物多样性的影响。
物种丰富度与本地生态系统的稳定性有密切联系, 同时也是影响生态系统生产力的重要因素, 其分布格局及驱动机制一直是生态学研究的核心问题之一[7–8]。在城市生物多样性研究中, 由于动态的城市相关数据较难获取, 因此城乡梯度(urban-rural gradient)往往作为“空间替代时间”的有效研究方法, 并应用于多种类群的研究中[9]。此外, 城市扩张往往伴随大量不透水表面的增加, 从乡村到城市中心, 不透水面占比逐渐增加, 因此不透水面占比可作为城市化强度的良好定量指标[10–11]。有关城乡梯度上物种丰富度分布格局及其驱动机制, 已有大量的假说[12–14], 较为常见的是干扰增长假说(increasing disturbance hypothesis)[13]及中度干扰假说[14](interme-diate disturbance hypothesis)。干扰增长假说预测物种丰富度随城市化干扰程度的增加将呈现单调递减的趋势, 中度干扰假说则认为物种丰富度将在中度干扰情况下达到最高。目前, 学界对城市中生物多样性分布格局仍然存在较大争议。Mcgill 等[15]对不同生物多样性在多尺度上的变化趋势进行总结, 发现物种丰富度在全球、生物地理区域、聚合群落及本地等 4 个尺度上均存在下降趋势。他们提出, 以往的研究对生物多样性类型及研究尺度缺乏关注是导致目前众说纷坛的主要原因。
我国是全球 12 个生物多样性大国之一, 但有关城市生物多样性的研究起步较晚, 相关研究集中于城市植物多样性, 且大多基于单个城市或单个区域[16], 缺少有关城市中哺乳动物丰富度的多空间尺度研究。中国哺乳动物丰富度分布格局与多种环境变量间存在显著的相关性, 方精云等[17]和王志恒等[18]从能量假说等角度进行解释, 但目前缺乏基于城乡梯度方法分析城市化影响的相关研究。
华东地区的长江三角洲(简称“长三角”)城市群是我国经济发展最快的地区, 面临着严重的生态压力[19], 研究该地区城市化对哺乳动物丰富度的影响, 对实现区域可持续发展及生态安全目标, 保护全国生物多样性有重要意义。本文基于城乡梯度方法, 研究长三角城市群哺乳动物丰富度沿城乡梯度的分布格局以及城市化、气候和生境特点等因素的影响。以华东地区作为背景地理区域, 通过对比分析城市群及其背景地理区域哺乳动物丰富度的分布格局及驱动因素, 分析城市化及背景气候对哺乳动物丰富度的贡献。
中国疆域辽阔, 地形复杂, 气候多样, 共有 58科 248 属 693 种哺乳动物[20]。华东地区位于我国东部, 属于亚热带湿润性季风气候和温带季风气侯, 该地区的长三角城市群(32°34′N ~29°20′N, 115°46′ E ~123°25′E)属于国家级城市群, 是我国经济增长最迅速、城市化水平最高的地区之一[19,21]。基于世界气候数据网站 (http://www.worldclim.org)提供的全球气候栅格数据计算得知, 长三角城市群的年平均气温为 15.59℃, 年平均降水量为 1248.8mm。依据相关统计年鉴数据[22–23], 2013 年长三角城市群的总市区人口为 7883.2 万, 城区人口为 5929.18 万, 城镇化率高达 75.2% (当年全国城镇化率为 54.1%), 人均 GDP 为 71440 元。2015 年, 长三角城市群的林地面积为 31683km2(占比为 28.70%), 草地面积为1387km2 (占比为 1.26%), 耕地面积为 50538km2(占比为 45.78%), 水域面积为 10671km2(占比为9.67%), 城镇居民用地面积为 16105km2(占比为14.59%)[24]。
哺乳动物丰富度数据来自国际自然保护联盟 (IUCN) 和哥伦比亚大学国际地球科学信息网络中心 (CIESIN) 共同汇编的全球哺乳动物丰富度网格数据集(https://doi.org/10.7927/H4N014G5), 该数据采集于 2008 年, 空间分辨率为 1 km²。
全球气候栅格数据来源于世界气候数据网站(http://www.worldclim.org), 空间分辨率约为 1km², 包含 1970—2000 年最低、平均和最高温度以及降水的平均气候数据。本文采用以下气候变量: 年平均温度(annual mean temperature, AMT)、气温日变幅(mean diurnal range, MDR)、最热月份最高温度(max temperature of warmest month, MTWM)、最冷月份最低温度(min temperature of coldest month, MTCM), 年平均降水(annual precipitation, AP)和最干旱月份降水(precipitation of driest month, PDM)。
生境数据来自空间分辨率为 30m 的 2000 年中国土地利用状况遥感监测数据 (CNLUCC), 由中国科学院资源环境科学数据中心(RESDC)根据 Landsat TM/ETM 遥感影像, 结合人工目视解译生成 (http:// www.resdc.cn)。本文将原始土地利用类型重新划分为六大类: 林地、草地、湿地、耕地、不透水表面和其他用地。利用软件 ArcMap 10.2 对土地数据进行重采样操作, 提取每 1km2 网格内 6 类用地的面积占比 (0~1)。计算各网格内的栖息地多样性指数(Shannon diversity index, SHDI)[25–26], 用来反映景观的丰富度和均匀度, 计算公式为
(1)其中, N 代表栖息地类型的数量, pi代表每 1 km²网格中栖息地类型 i 的占比。当栖息地类型数量增加和/或不同栖息地类型分布趋向均匀时, SHDI数值也随之增大。当 SHDI=0 时, 表示网格中只存在一种栖息地类型。
夜间灯光数据(nighttime light, NL)可较好地反映社会的人口和经济情况[27–28]。本文将夜间灯光作为社会经济因子的定量指标, 反映城市化的间接影响。中国的夜间灯光数据来源于全球 DMSP-OLS夜间灯光栅格数据集, 该数据由美国国家海洋与大气管理局的国家地球物理数据中心(NOAA-NGDC)进行处理和分发。这些夜间卫星图像记录从地球表面观察到的光发射量, 本文使用的是由 DMSP 卫星F15 在 2000 年收集的稳定光数据。
将每 1km² 网格内不透水面的面积占比作为该网格的城市化强度(urbanization intensity, UI) 定量指标, 并以每增加 1% 的不透水表面占比作为节点, 将从 0~1 的城乡梯度划分为 101 个区间。使用皮尔逊相关分析方法, 分析 101 个区间内哺乳动物丰富度与气候变量、栖息地变量、城市化强度及社会经济因子等 14 个影响因素间的单因素相关关系。通过逐步回归分析, 考察多因素综合作用下, 影响长三角城市群及华东地区哺乳动物丰富度沿城乡梯度变化的主导驱动因子。统计和分析过程均用软件 R 3.4.1 完成。
长三角城市群的哺乳类丰富度平均值为 51, 主要在 45~60 之间。华东地区的哺乳动物丰富度的平均值为 53, 且主要在 45~60 之间。研究表明, 我国哺乳动物丰富度高值区集中在西南地区和中南地区, 哺乳动物丰富度整体上呈从低纬向高纬地区递减趋势[17]。本文结果与上述研究相符, 长三角城市群及华东地区的哺乳动物丰富度整体上呈现“北少南多” 的空间分布情况。
长三角城市群的哺乳动物丰富度沿城乡梯度的空间分布格局与其背景地理区域尺度的分布格局相似, 均呈现水平 “S” 分布模式(图 1)。长三角城市群和华东地区的哺乳动物丰富度均在城市化强度为 0 时达到最高, 但在该城市化强度区间内, 长三角哺乳动物丰富度的中位数低于华东地区。从城市化强度为 0 到低度城市化地区(0≤UI≤0.25), 长三角城市群和华东地区哺乳动物丰富度均呈显著下降趋势; 从低度城市化到高度城市化地区, 哺乳动物丰富度缓慢上升。当城市化强度超过 0.90 时, 两个空间尺度的哺乳动物丰富度均明显下跌。
根据皮尔逊相关分析结果(表 1), 在长三角城市群尺度, 除最热月份最高温度外, 其余气候变量均与长三角城市群哺乳动物丰富度显著相关(P< 0.05)。气温日变幅与哺乳动物丰富度显著正相关, 年平均温度、最冷月份最低温度、年平均降水及最干旱月份降水则与哺乳动物丰富度显著负相关。在华东地区, 所有气候变量均与哺乳动物丰富度显著相关, 且与对应变量在城市群尺度各自的相关关系恰好相反。
长三角城市群的哺乳动物丰富度与城市化强度、林地面积占比以及夜间灯光显著正相关, 与耕地面积占比以及栖息地多样性指数显著负相关(P< 0.05)。在华东地区, 哺乳动物丰富度与城市化强度及栖息地多样性指数显著负相关, 但与林地、草地及其他用地面积占比显著正相关(P<0.05)。

菱形点为丰富度平均值
图1 华东地区及长三角城市群哺乳动物沿城乡梯度空间分布箱线图
Fig. 1 Boxplots of mammal richness’s spatial distribution pattern along the urban-rural gradient in East China and Yangtze River Delta urban agglomeration
表1 华东地区及长三角城市群皮尔逊相关分析及逐步回归结果
Table 1 Results of Pearson correlation analysis and stepwise regression analysis in East China and Yangtze River Delta urban agglomeration

自变量华东地区长三角城市群 皮尔逊相关系数逐步回归系数逐步回归R2/%皮尔逊相关系数逐步回归系数逐步回归R2/% 年平均温度0.485***–0.286** 气温日变幅–0.458***0.197* 最热月最高温–0.521***0.2010.50.141 最冷月最低温0.621***0.0390.5–0.644***–0.08141.4 年平均降水0.934***0.41487.2–0.640***–0.3553.2 最干旱月份降水0.700***–0.534*** 城市化强度–0.188*–2.7780.20.238** 林地面积占比0.825***–0.3540.60.519***0.65626 草地面积占比0.533***0.1128.5–0.030 湿地面积占比0.5320.027 耕地面积占比–0.143–2.6660.3–0.518***–0.37010.5 其他用地面积占比0.585***0.140 栖息地多样性指数–0.267**–0.376*** 夜间灯光–0.0860.279** 逐步回归总解释度97.781.1
注: * P < 0.05, ** P < 0.01, *** P < 0.001。
当综合考虑各变量间的相互影响时, 两个空间尺度上的主要解释变量存在共性, 即温度和降水等气候变量成为影响哺乳动物丰富度的主要解释因子。在华东地区, 年平均降水对哺乳动物丰富度沿城乡梯度变化的解释度最高(R2=87.2%, 表 1), 在长三角城市群, 解释度最高的气候变量为最冷月最低温(41.1%)。图 2 和 3 为各变量的平均值沿城乡梯度的变化趋势, 可以发现, 华东地区和长三角城市群的主导解释变量沿城乡梯度的各自变化趋势与哺乳动物丰富度在以上两个空间尺度沿城乡梯度的分布模式十分相似。此外, 尽管气候变量在两个空间尺度上均为主要解释因子, 但在华东地区, 城市化强度对哺乳动物丰富度存在负面影响 (0.2%)。同时, 林地、耕地面积占比在两个空间尺度上均被纳入最终的逐步回归方程, 且城市化相关因素在城市群尺度上的解释率相较背景区域尺度更高——长三角城市群的林地面积占比对哺乳动物丰富度存在正面影响(26%), 耕地面积占比则存在负面影响(10.5%)。
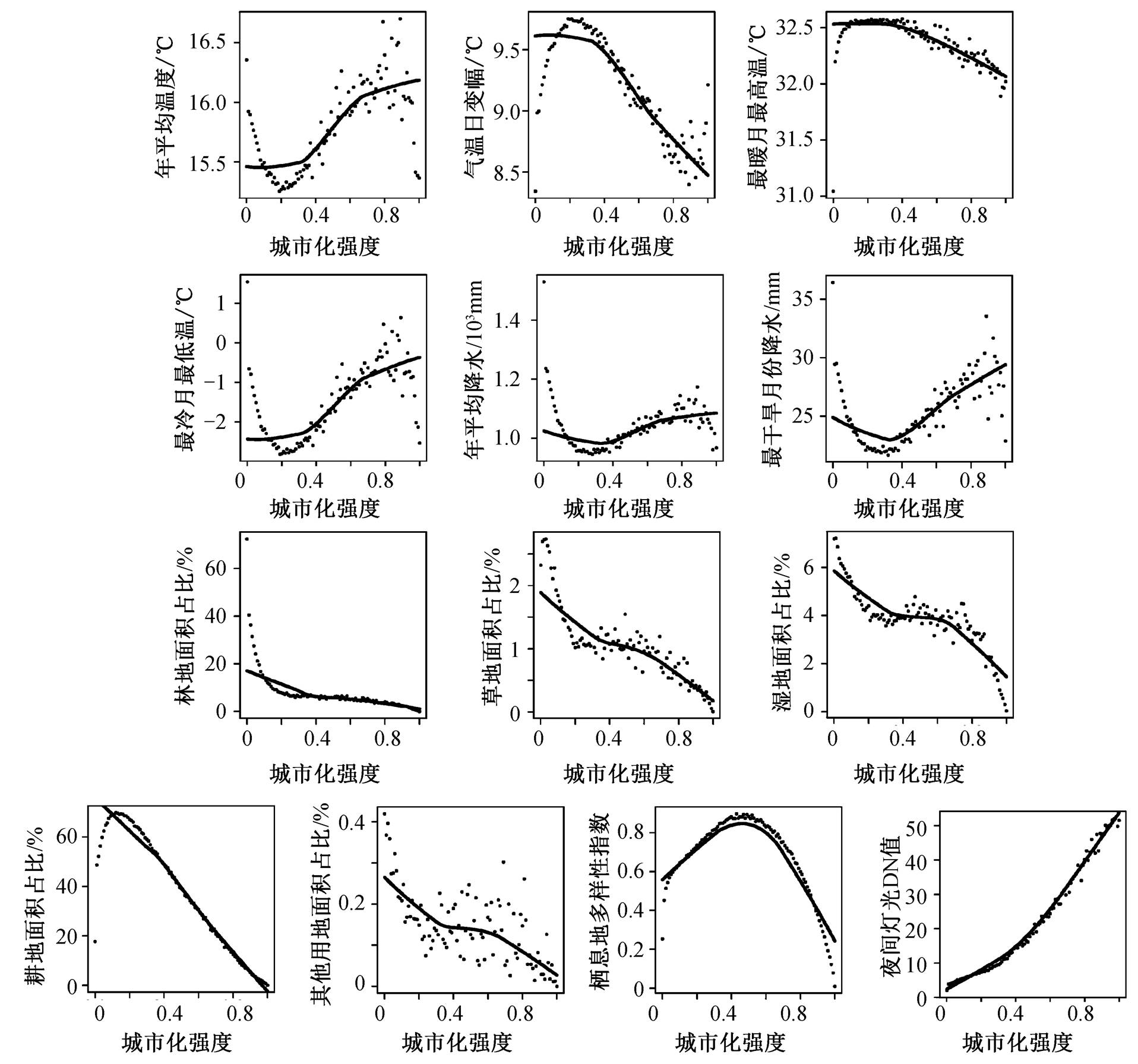
图2 华东地区各自变量平均值沿城乡梯度分布散点图
Fig. 2 Scatterplots of the mean values of each independent variables’ distribution along the urban-rural gradient in East China
本文研究结果表明, 在两个空间尺度下, 哺乳动物丰富度沿城乡梯度的分布格局与干扰假说和中度干扰假说不一致: 随着城市化强度的增加, 长三角城市群及华东地区哺乳动物丰富度并非呈现单调递减或倒“U”型曲线, 而是呈现出水平的“S”型分布。这个结果是合理的, 因为以往研究对城乡梯度的划分较为粗糙, 只进行定性的划分或简单地划分为“低、中、高”3 个梯度[9], 在复杂交错的城市环境中, 城乡梯度上哺乳动物丰富度的细微变化容易被粗略的梯度划分掩盖。
若仅考虑城乡梯度上哺乳动物丰富度分布曲线的起点和终点, 则无论是背景地理区域尺度还是城市群尺度, 哺乳动物丰富度均值都在城市化强度为0 时达到最高, 高度城市化地区的丰富度则相对较低, 与增长干扰假说相符。值得注意的是, 在高度城市化地区, 哺乳动物丰富呈显著下降趋势, 该结果与 Mckinney[9]基于 105 篇文献的 Meta 分析结果一致, 即对不同的脊椎动物而言, 高度城市化均会导致其丰富度下降。
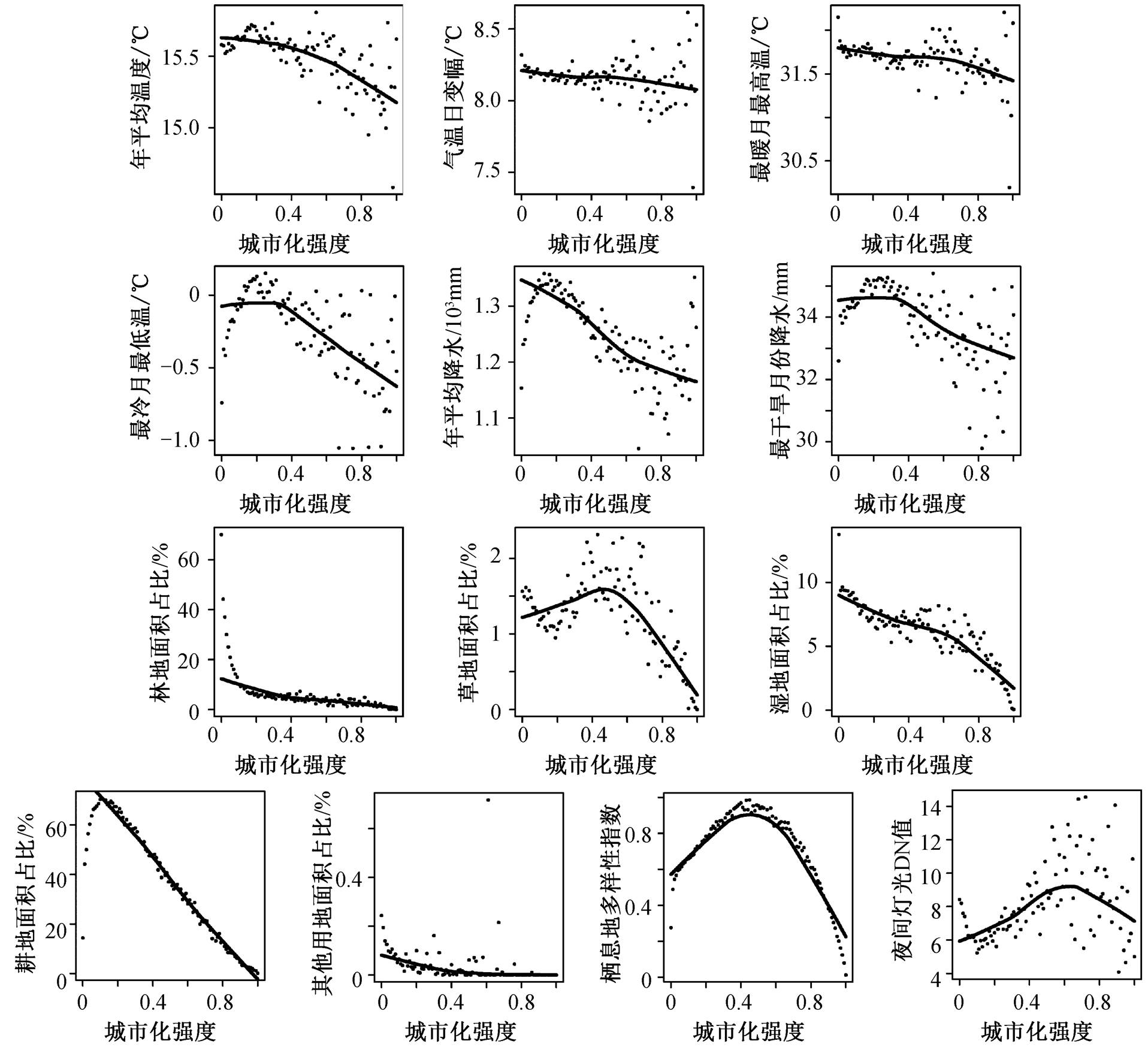
图3 长三角城市群各自变量平均值沿城乡梯度分布散点图
Fig. 3 Scatterplots of the mean values of each independent variables’ distribution along the urban-rural gradient in Yangtze River Delta urban agglomeration
从低度城市化地区到中度城市化地区, 长三角城市群哺乳动物丰富度存在缓慢上升趋势。长三角城市群的栖息地多样性指数在城市化强度为 0.5 时达到最高(图 3)。根据中度干扰假说, 哺乳动物丰富度应与栖息地多样性指数存在相似的分布模式, 然而皮尔逊相关分析结果表明, 两者间呈显著负相关关系(r = ‒0.376, P<0.001, 表 1)。此外, 中度干扰假说未能解释从城市化强度为 0 到低度城市化地区(0≤ UI≤ 0.25)时, 华东地区和长三角哺乳动物丰富度的明显下降趋势。
在背景地理区域和城市群尺度上, 温度、降水等气候变量与哺乳动物丰富度均存在显著相关关系(p<0.05, 表 1)。其中, 华东地区的年平均降水量与哺乳动物丰富度之间强正相关(r=0.934, P<0.001), 且是影响哺乳动物丰富度的最主导解释变量(R2= 87.2%); 最冷月份最低温度同样与华东地区的哺乳动物丰富度显著正相关, 即便在多因素综合作用时, 也是重要解释变量之一(r=0.621, P<0.001, R2= 0.05%)。在方精云等[17]基于 900 个 100km×100km中国物种网格数据进行的有关中国陆生哺乳动物丰富度格局与多种环境变量间关系的研究中, 同样发现年平均降水量以及最冷月份最低温度与哺乳动物丰富度存在强正相关关系(AP: r=0.686, P<0.05; MTCM: r=0.627, P<0.05)。以上结果均符合环境热量假说(ambient-energy hypothesis)[29]和水分–能量假说(water-energy hypothesis)[30]。
环境热量假说认为能量对大尺度物种丰富度格局的形成起主导作用, 能量主要是通过影响物种的体温调节、生存繁衍及分化等生理过程来影响种群中的物种数量[29]。水分–能量假说则认为水分以及热量共同影响大尺度的物种多样性格局[30]。结合Currie[29]对环境热量假说的总结, 可认为良好的水热环境有利于更多物种生存[31]。作为恒温动物, 哺乳动物需要通过消耗能量来维持稳定的体温, 处在寒冷的环境中会加大哺乳动物用于体温调节的能量消耗[32]。因此, 一个水热条件较好的环境, 有利于哺乳动物减少内部能量消耗, 从而将更多的能量用于物种繁衍和分化, 提高物种丰富度。华东地区的年平均降水量沿城乡梯度同样呈现大致水平的“S”型分布, 此外, 华东地区城市化强度为 0 的地区, 其年平均降水量及最冷月份最低温度均高于存在城市化干扰的地区(UI>0, 图 2), 同时还具有相对较高的年平均气温, 这也能部分地解释华东地区哺乳动物丰富度沿城乡梯度呈水平的 “S” 型分布格局以及为何其丰富度在城市化强度为 0 的地区达到最高。
本文研究结果表明, 不同空间尺度上哺乳动物丰富度受城市化和气候因素的影响程度不同。背景区域尺度上, 气候变量对华东地区哺乳动物丰富度总解释率接近 90%, 但在长三角城市群, 气候变量的总解释率仅超过 40%, 而城市化相关因素的总解释率则由华东地区的 9.6%增大至 36.5%。因此, 城市化对哺乳动物丰富度影响存在尺度依赖性, 在自然地理区等较大空间尺度上, 气候变量对哺乳动物丰富度格局的影响更显著。
尽管在背景区域尺度, 背景气候是影响哺乳动物丰富度格局的主导因素, 但随着空间尺度缩小至城市群, 其内部的温度、降水等气候条件随着城市化强度增加变化幅度均小于华东地区(图 2 和 3), 城市化对哺乳动物丰富度的直接和间接影响也逐渐凸显。城市化能通过改变栖息地类型, 尤其是通过影响城乡梯度上林地和耕地的面积占比, 直接影响哺乳动物丰富度。一方面, 长三角城市群的哺乳动物丰富度与林地面积占比之间显著正相关(r=–0.519, P<0.001, 表 1), 且其对长三角城市群哺乳动物丰富度变化的解释率(R2=26%)仅次于最冷月份最低温度(R2 = 41.4%); 另一方面, 长三角城市群哺乳动物丰富度与耕地面积占比显著负相关(r=–0.518, P< 0.001, R2=10.5%)。此外, 城市化强度在华东地区与哺乳动物丰富度呈弱负相关关系(r=–0.188, P< 0.05, R2=0.2%), 但在长三角城市群, 城市化强度及夜间灯光均与哺乳动物丰富度弱正相关 (UI:r = 0.238, P<0.01; NL: r=0.279, P<0.01), 可能与城市化的间接影响有关。
原始天然林地对城市中哺乳动物的种群生存繁衍有至关重要的作用。城市化的发展往往伴随着栖息地破碎化和生境异质性, 从低度城市化地区到城市中心, 越来越多的自然植被被不透水面替代, 大面积自然林地逐渐破碎化, 乃至消失[7,11]。从无城市化干扰到低度城市化地区 (0≤UI≤0.25), 长三角城市群的平均林地面积占比大幅度下降(图 3), 这是导致哺乳动物丰富度在此区间迅速下跌的原因之一。根据种面积理论(species-area theory), 面积更大的斑块更容易出现更多的物种[33]。不同于能从多样化栖息地获益的部分植物(如广幅种或生存在斑块交界处的植物), 体形较大、巢区范围广的哺乳动物通常需要更大面积的栖息地来维持种群的生存[34–35], 更容易受到栖息地破碎化的负面影响。所以, 对长三角城市群的哺乳动物而言, 城市化带来的负面影响可能大于生境异质性可能带来的正面影响。本研究中, 哺乳动物丰富度在城市群尺度及背景地理区域尺度均与栖息地多样性指数呈显著负相关关系(表 1), 同样证明了以上结论。此外, 研究表明, 城市密集的交通活动是导致城市地区哺乳动物死亡率较高的原因之一[36], 林地则为城市中野生动物提供了一个远离人类活动干扰的庇护所, 因此保护天然林地、修复城市中残存自然林地斑块, 对保护城市中的哺乳动物多样性有重要意义。
农业活动同样是对地表改变剧烈的人类活动之一, 包括将非农用地转化农业用地以及增强原有农业用地的利用强度。1961—2000 年, 全世界发展中国家的耕地总面积增加 20%, 预计到 2050 年全球粮食需求将增加一倍以上[37]。本研究中, 从城市化强度为 0 到低城市化地区 (0≤UI≤0.25), 长三角城市群哺乳动物丰富度呈显著下降趋势, 耕地平均面积占比迅速增加 (图 3), 与我国乡村地区一直是农业活动重点开展地区的国情有关。根据陈肖飞等[38]对1990 年以来长江三角洲耕地资源变化的研究结果, 长三角地区的耕地实际利用强度整体上呈现“北强南弱”的分布格局, 与长三角城市群哺乳动物“北少南多”的整体空间分布格局类似。上述结果表明, 耕地面积的增加可能是导致长三角城市群的哺乳动物丰富度减少的另一重要原因。
以前的研究发现, 农业活动对生物多样性也存在负面影响。Green 等[37]基于国际鸟类保护组织的世界鸟类数据库, 研究农业活动对全球 1923 种受威胁(threatened)和接近威胁(near-threatened)鸟类的影响, 表明农业活动是目前和未来影响鸟类生存的主要威胁, 尤其对发展中国家的鸟类而言。Kehoe等[39]将 13 个表征农业土地利用强度(LUI)的指标分别与哺乳动物、鸟类和两栖动物的特有丰富度(en-demism richness)地图叠加, 研究目前全球高 LUI 地区与高生物多样性地区的空间重合情况, 结果表明, 鸟类和哺乳动物的空间分布格局惊人地相似, 即约 80%同时具备高 LUI 和高鸟类特有丰富度的地区, 其哺乳动物特有丰富度也较高。此外, 许多既是高 LUI, 也具有高物种特有丰富度的地区(例如南美、中国和东部非洲)未列入公认的生物多样性热点区域。随着农业集约化进程的加快, 许多目前LUI 较低, 但物种特有丰富度较高的地区(主要位于中美洲、非洲东部和东南亚), 其生物多样性可能正处于危险之中。中国虽然是一个农业大国, 但人均耕地面积远低于国际水平, 近年来经济发展和耕地资源间的矛盾也愈演愈烈, 而长三角城市群恰好是我国最重要的商品粮基地之一。因此在未来城市化建设过程中, 还需要更多地关注我国乡村地区的生态友好型农业建设, 从而实现经济发展、农业生产与当地生物多样性保护的平衡。
城市的生态足迹往往远远超越其实际地理边界[40], 除城市化对哺乳动物丰富度的直接影响外, 城市化的间接影响(如城市热压力、环境污染、人为景观管控和社会经济活动等)更值得学者们给予关注[41–42]。Kinzig 等[43]检验了社会经济和文化因素对鸟类和植物物种丰富度的影响, 发现具有不同社会经济和文化特征居民区的相关物种丰富度沿城乡梯度存在显著的变化, 表明城市化可以通过社会经济活动和影响人口密度而间接地影响城市生物多样性。此外, DMSP/OLS 夜间影像的亮度与区域GDP间存在显著的相关关系, 因此夜间灯光通常可作为社会经济活动的良好替代指标[44]。本研究中, 背景区域尺度上夜间灯光与哺乳动物丰富度并无显著的相关关系, 但在城市群尺度上, 夜间灯光与哺乳动物丰富度间存在弱正相关关系(r=0.279, P<0.001)。Jia等[45]基于概念框架和美国 377 个大都市区的遥感数据, 研究城市化对植被生长的直接和间接影响, 发现在 80%的城市网格中, 城市化的间接影响明显促进植被的生长。以上结果表明, 城市化对生物多样性的间接影响也许并不都是负面的。
Jin 等[46]利用生态足迹模型, 对生态压力指数和生态效率(ecological efficiency, EE)进行定量的描述, 描述自然资源可持续性和利用效率的现状, 分析我国 30 个省份经济集聚状况(economic agglo-meration, EA)与生态效率之间的关系, 结果表明, 除青海外, 其他省份均面临生态超负荷情况, 其中上海、天津和北京的生态超负荷最为严重。长三角城市群所在的上海、江苏和浙江均为高 EA 和高EE 地区, 面临严峻的生态压力。Jin 等[46]指出, 经济集聚与生态效率之间存在正相关关系, 促进经济集聚是提高自然资源利用效率的有效方法。作为我国综合发育水平最高的地区, 长三角城市群是我国经济增长的重要引擎, 研究其社会经济活动与物种丰富度间的联系, 有助于我国保持经济增长的同时, 实现区域可持续发展及生态安全目标。
城市化是人地交互作用的复杂地理过程, 存在多种因素的相互作用。城市的年龄、结构和发展路径等对生物多样性均存在影响[47]。不同生理结构和生活习性的物种对环境变量的响应也存在差异。因此, 凭借单个类群或单个城市群的研究结果, 难以全面地概括城市地区物种丰富度复杂多样的分布格局及影响因素。本研究是基于定量的和细化的城乡梯度, 探讨城市化及气候背景等多种因素对长三角城市群哺乳动物丰富度的相对影响, 后续研究中, 应针对不同类群的物种, 结合多个空间尺度和不同城市群, 区分城市化与背景气候对生物多样性的影响, 并对城市化对生物多样性的间接影响给予更多的关注。
参考文献
[1] Botzat A, Fischer L K, Kowarik I. Unexploited op-portunities in understanding liveable and biodiverse cities. A review on urban biodiversity perception and valuation. Global Environmental Change, 2016, 39: 220–233
[2] Grimm N B, Faeth S H, Golubiewski N E, et al. Global change and the ecology of cities. Science, 2008, 319: 756–760
[3] Seto K C, Fragkias M, Güneralp B, et al. A Meta-analysis of global urban land expansion. PLOS ONE, 2011, 6(8): e23777
[4] Seto K C, Burak G, Hutyra L R. Global forecasts of urban expansion to 2030 and direct impacts on bio-diversity and carbon pools. Proceedings of the Na-tional Academy of Sciences of the United States of America, 2012, 109(40): 16083–16088
[5] Magle S B, Hunt V M, Vernon M, et al. Urban wildlife research: past, present, and future. Biological Conservation, 2012, 155: 23–32
[6] Lahr E C, Dunn R R, Frank S D. Getting ahead of the curve: cities as surrogates for global change. Procee-dings of the Royal Society B: Biological Sciences, 2018, 285: 20180643
[7] Mckinney M L. Urbanization as a major cause of biotic homogenization. Biological Conservation, 2006, 127(3): 247–260
[8] Mcgill B J. Towards a unification of unified theories of biodiversity. Ecology Letters, 2010, 13(5): 627–642
[9] Mckinney M L. Effects of urbanization on species richness: a review of plants and animals. Urban Eco-systems, 2008, 11(2): 161–176
[10] Peng J, Hu Y N, Liu Y X, et al. A new approach for urban-rural fringe identification: integrating imper-vious surface area and spatial continuous wavelet transform. Landscape & Urban Planning, 2018, 175: 72–79
[11] Mckinney M L. Urbanization, biodiversity, and con-servation. Bioscience, 2002, 52(10): 883–890
[12] Marquet P A, Allen A P, Brown J H, et al. On theory in ecology. Bioscience, 2014, 64(8): 701–710
[13] Gray J S. Effects of environmental stress on species rich assemblages. Biological Journal of the Linnean Society, 1989, 37(1/2): 19–32
[14] Connell J H. Diversity in tropical rain forests and coral reefs. Science, 1978, 199: 1302–1310
[15] Mcgill B J, Dornelas M, Gotelli N J, et al. Fifteen forms of biodiversity trend in the Anthropocene. Trends in Ecology & Evolution, 2015, 30(2): 104–113
[16] 毛齐正, 马克明, 邬建国, 等. 城市生物多样性分布格局研究进展. 生态学报, 2013, 33(4): 1051–1064
[17] 方精云, 林鑫, 王志恒, 等. 中国陆栖哺乳动物物种丰富度的地理格局及其与环境因子的关系. 生物多样性, 2009, 17(6): 652–663
[18] 王志恒, 唐志尧, 方精云. 物种多样性地理格局的能量假说. 生物多样性, 2009, 17(6): 613–624
[19] 王祥荣, 樊正球, 谢玉静, 等.城市群生态安全保障关键技术研究与集成示范——以长三角城市群为例. 生态学报, 2016, 36(22): 7114–7118
[20] 蒋志刚, 马勇, 吴毅, 等. 中国哺乳动物多样性. 生物多样性, 2015, 23(3): 351–364
[21] 黄金川, 陈守强. 中国城市群等级类型综合划分. 地理科学进展, 2015, 34(3): 290–301
[22] 长江三角洲城市年鉴编辑委员会. 长江三角洲城市年鉴. 北京: 中国文史出版社, 2014
[23] 中国城市建设统计年鉴编辑委员会. 中国城市建设统计年鉴. 北京: 中国计划出版社, 2013
[24] 陈书林, 虞宋杰. 长三角地区 1985~2015 年土地利用变化及其驱动力分析. 林业经济, 2019(11): 49–54
[25] Shannon C E, Weaver W, Wiener N. The mathematical theory of communication. Urbana: University of Illi-nois Press, 1949
[26] Nagendra H. Opposite trends in response for the Shannon and Simpson indices of landscape diversity. Applied Geography, 2002, 22(2): 175–186
[27] Nordhaus W, Chen X. A sharper image? Estimates of the precision of nighttime lights as a proxy for eco-nomic statistics. Journal of Economic Geography, 2015, 15(1): 217–246
[28] Tang L, Cheng H, Qu G. Estimating provincial ec-onomic development level of China using DMSP/OLS nighttime light satellite imagery. Advanced Materials Research, 2013, 807/808/809: 1903–1908
[29] Currie D J. Energy and large-scale patterns of animal-and plant-species richness. The American Naturalist, 1991, 137(1): 27–49
[30] O’Brien E M. Climatic gradients of woody plant species richness towards an explianation based on analysis of Southern Africa’s woody flora. Journal of Biogeography, 1993, 20(2): 188–198
[31] Kleidon A, Mooney H A. A global distribution of biodiversity inferred from climatic constraints: results from a process-based modelling study. Global Change Biology, 2010, 6(5): 507–523
[32] Greenville A C, Wardle G M, Dickman C R. Extreme climatic events drive mammal irruptions: regression analysis of 100-year trends in desert rainfall and tem-perature. Ecology & Evolution, 2012, 2(11): 2645–2658
[33] Palmer M W, White P S. Scale dependence and the species-area relationship. American Naturalist, 1994, 144(5): 717–740
[34] Noss R F, Quigley H B, Hornocker M G, et al. Conservation biology and carnivore conservation in the rocky mountains. Conservation Biology, 1996, 10 (4): 949–963
[35] Šálek M, Drahníková L, Tkadlec E. Changes in home range sizes and population densities of carnivore species along the natural to urban habitat gradient. Mammal Review, 2015, 45(1): 1–14
[36] Rytwinski T, Fahrig L. The impacts of roads and traffic on terrestrial animal populations. Chichester: John Wiley & Sons, Ltd, 2015
[37] Green R E, Cornell S J, Scharlemann J R P W, et al. Farming and the fate of wild nature. Science, 2005, 307: 550–555
[38] 陈肖飞, 姚士谋, 张落成. 1990 年以来长江三角洲耕地资源变化及驱动因子研究. 长江流域资源与环境, 2015, 24(9): 1521–1527
[39] Kehoe L, Kuemmerle T, Carsten M, et al. Global patterns of agricultural land-use intensity and verte-brate diversity. Diversity & Distributions, 2015, 21 (11): 1308–1318
[40] Ree W E. Ecological footprints and appropriated carrying capacity: what urban economics leaves out. Environment and Urbanization, 1992, 4(2): 121–130
[41] Aronson M F J, Nilon C H, Lepczyk C A, et al. Hierarchical filters determine community assembly of urban species pools. Ecology, 2016, 97(11): 2952–2963
[42] Mcdonald R I, Mansur A V, Ascensão F, et al. Research gaps in knowledge of the impact of urban growth on biodiversity. Nature Sustainability, 2019, 3: 16–24
[43] Kinzig A P, Warren P, Martin C, et al. The effects of human socioeconomic status and cultural characteris-tics on urban patterns of biodiversity. Ecology and Society, 2005, 10(1): no. 23
[44] Dai Z, Hu Y, Zhao G. The suitability of different nighttime light data for GDP estimation at different spatial scales and regional levels. Sustainability, 2017, 9(2): 305–320
[45] Jia W, Zhao S, Liu S. Vegetation growth enhancement in urban environments of the Conterminous United States. Global Change Biology, 2018, 24: 4084–4094
[46] Jin X, Li X, Feng Z, et al. Linking ecological efficiency and the economic agglomeration of China based on the ecological footprint and nighttime light data. Ecological Indicators, 2020, 111: 106035
[47] Saari S, Richter S, Higgins M, et al. Urbanization is not associated with increased abundance or decreased richness of terrestrial animals-dissecting the literature through meta-analysis. Urban Ecosystems, 2016, 19 (3): 1–14
Urbanization Effects on Mammal Richness: A Case Study of Yangtze River Delta Urban Agglomeration
Abstract Taking the fraction of impervious surface area as a quantitative indicator of urbanization intensity (UI), using the global mammal richness girds data set and combined with climatic variables and habitat characteristics, this paper studied the spatial distribution pattern of mammal richness along the urban-rural gradient in Yangtze River Delta urban agglomeration. The contribution of urbanization and background climate were analyzed by comparing the richness distribution pattern and driving forces of Yangtze River Delta urban agglomeration and the geographic region (East China) where it located. The results were as follows. 1) There was a horizontal “S” distribution pattern of mammal richness along the urban-rural gradient at both spatial scales. 2) In background geographic region, climatic variables (e.g, annual precipitation and min temperature of coldest month) played a dominant role in richness distribution (R2= 88.2%). 3) There was a negative correlation between UI and richness in East China while a weakly positive correlation in Yangtze River Delta (P<0.001); woodland had positive impacts on richness, while arable land had negtive impacts. The urbanization effects on mammal richness were more pronounced in Yangtze River Delta. Future research should pay more attention to how to disentangle the impacts of urbanization from background climate on biodiversity.
Key words urban-rural gradient; mammals; biodiversity; impervious surface; urbanization intensity; back-ground climate
doi: 10.13209/j.0479-8023.2021.036
收稿日期: 2020-04-11;
修回日期: 2020-05-22