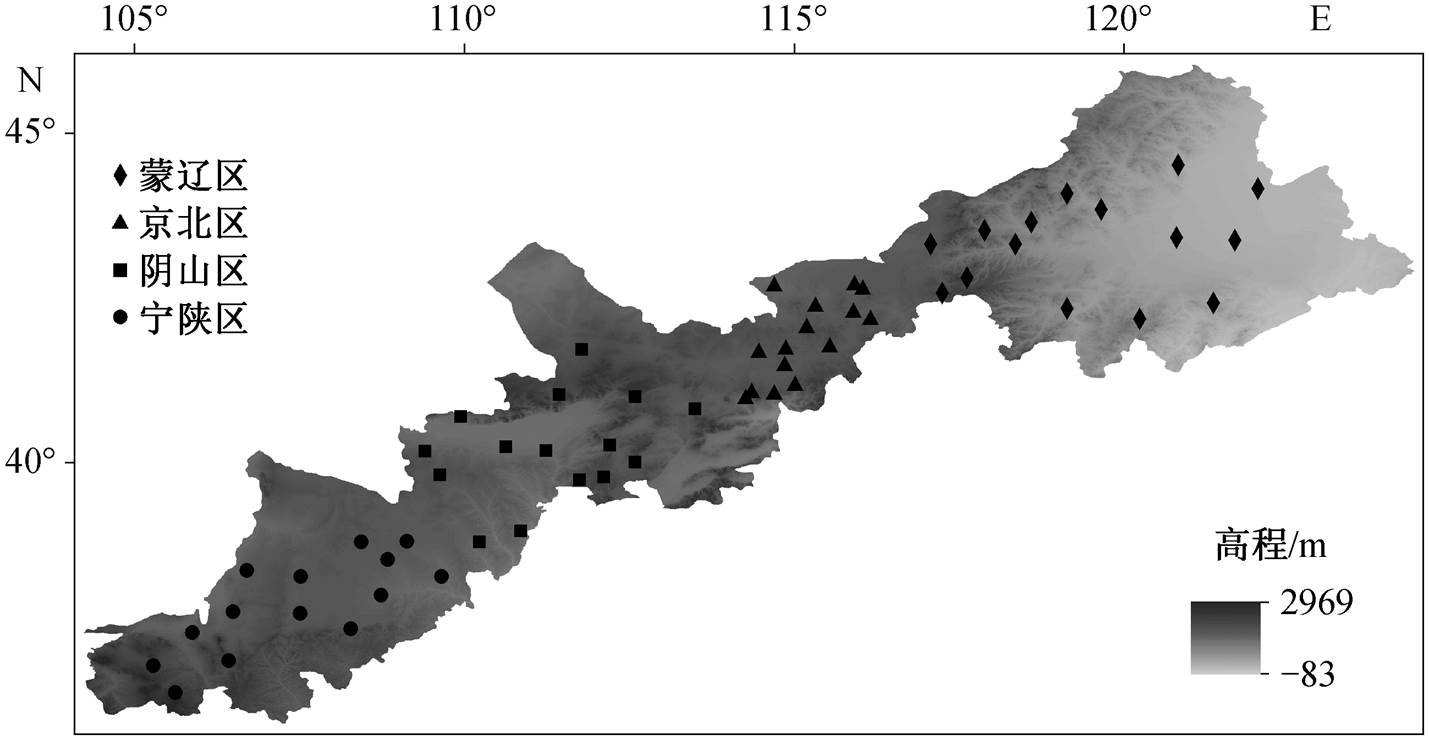
图1 样点位置
Fig. 1 Locations of the sampling sites
摘要 为探究我国北方农牧交错带草地表层(0~10cm)和次表层(10~20cm)土壤微生物量碳的空间格局及其环境驱动力, 选择蒙辽平原、京北坝上、阴山北麓和宁陕黄土高原 4 个区域总计 456 个土壤样品(两个土层, 57个样点, 每个样点 4 个样方)进行调查。结果表明, 表层与次表层土壤微生物量碳的空间格局具有一致性, 均随纬度增加而增加, 随经度和海拔增加无显著变化。随着草地退化程度加剧, 次表层土壤微生物量碳的降低幅度小于表层土壤。两层土壤 pH 值的差异随草地退化程度的加剧而缩小, 表层与次表层土壤微生物量碳的差异受土壤 pH 值调控, 两个土层的 pH 值差异越小, 微生物量碳的差异越小。气候、植被和土壤因素均会影响微生物量碳的空间变异, 其中土壤因素为区域尺度微生物量碳的主要驱动力, 表层土壤微生物量碳的主要影响因素为土壤总碳, 次表层土壤微生物量碳受土壤总氮的影响最大。在气候变化和人类活动影响加剧的背景下, 研究结果对预测区域尺度土壤微生物的响应规律及退化草地生态功能的维持和修复有重要意义。
关键词 微生物量碳(MBC); 土壤深度; 农牧交错带; 草地生态系统; 土壤pH
土壤微生物是生态系统的主要分解者[1], 直接参与碳固持[2]和养分循环[3-4]等一系列生态过程, 驱动陆地生态系统的生物地球化学循环[5]。土壤微生物量碳是土壤养分的活性部分[6], 也是土壤有机碳的重要组成[7-8], 对土壤有机质的转化和循环产生直接贡献[9], 影响植物生长[10]。土壤微生物量碳对环境变化十分敏感, 是评估土壤肥力和生态系统生产力的有效指标[11]。在区域尺度开展微生物量碳空间分布的研究, 有助于提升对全球变化背景下土壤碳循环的认识, 同时对微生物的生物地理学研究具有重要意义[6]。
目前, 对微生物量碳分布格局的研究主要关注表层土壤[12-14]。随着土壤深度增加, 土壤养分与理化性质均会发生显著的改变[15], 从而影响次表层土壤微生物量碳的分布格局。因此, 在区域尺度上, 次表层土壤与表层土壤微生物量碳的地理格局变化规律是否一致, 这一问题有待探究。研究表明, 气候、植被和土壤因素均会影响微生物量碳[13,16-17]。在干旱和半干旱草地开展的研究显示, 土壤因素(如土壤碳含量和土质等)是微生物量碳的主要驱动力[16]。Zhao 等[13]通过温带草地与高寒草地的整合分析, 发现气候因素对微生物量碳的影响更显著。彭晓茜等[17]对内蒙古温带草原的研究表明, 植被生物量也会对土壤微生物量碳产生一定程度的影响。但是, 次表层土壤微生物量碳的主要环境驱动力是否与表层土壤一致, 目前仍不清楚。
草地是分布最广泛的植被类型, 是陆地生态系统的重要组成部分[18]。随着人类活动和气候变化加剧, 草地退化已成为全球性问题[19]。在中国, 50%~60%的草地面临不同程度的退化[20]。以往对草地退化的研究集中在植被生产力、多样性和土壤肥力方面[21-22], 对土壤微生物量碳关注较少, 且集中在表层土壤[23]。通常, 表层土壤微生物量碳含量高于次表层土壤[13,17]。贺凤鹏等[23]在对内蒙古温带草地的研究中发现, 草地退化越严重, 表层土壤与次表层土壤的微生物量碳差异越小, 在极度退化的草地, 次表层土壤微生物量碳含量甚至高于表层土壤。这一规律对区域尺度上微生物量碳格局是否适用, 还有待验证。
我国北方农牧交错带位于干旱与半干旱气候的交汇带, 作为中东部地区重要的生态安全屏障和京津冀地区重要的水源涵养带, 对国家的生态安全具有重要意义[24]。近年来, 由于人类活动加剧, 农牧业过度经营, 该区域草地退化现象日益严重[24-25], 已成为又复杂又脆弱的生态系统[26]。深入理解该区域土壤微生物量碳的分布及驱动机制, 对维持区域生态系统功能具有现实意义。
基于上述背景, 本研究在我国北方农牧交错带4 个区域采集 456 个草地土壤样品(两个土层, 57 个样点, 每个样点 4 个样方), 探究区域尺度上表层与次表层土壤微生物量碳的空间分布格局及其环境驱动力的差异, 希望研究结果有助于预测全球变化及人类活动加剧背景下土壤微生物的响应规律, 为退化草地的修复提供科学依据。
研究区位于我国北方农牧交错带, 包括蒙辽平原(ML)、京北坝上(JB)、内蒙古阴山北麓(YS)和宁陕黄土高原(NS) 4 个区域, 属于东亚季风气候区与大陆干旱气候区的交错地带[27]。年平均气温为1.6~10.3℃, 年平均降水量为 245~513 mm。
结合气候、地貌、土壤基本理化性质和植被NDVI 数据, 采用分层取样的方法[28]进行采样, 57个样点的位置见图 1, 具体信息见表 1。
于 2017 年和 2018 年 7—8 月的植物生长最高峰季节进行土壤和植被样品采集。在每个样点随机选取 4 个 1m×1m 的样方, 将样方内地上部分植物从基部整齐地收割, 烘干称重后用于计算植被地上生物量碳。每个样方内, 用五点取样法分别采集 0~10和 10~20cm 两个深度土层的样品, 并将同层土壤样品混合均匀。土壤样品过 2mm 筛, 人工除去植物根系、植物残体、石块和动物等异物后装入封口袋, 及时放入有冰袋的保温盒中, 尽快运回实验室, 在 4℃条件下保存。
用氯仿熏蒸法[29]测定土壤微生物量碳: 称取两份 10.00 g 新鲜土壤样品于玻璃烧杯中, 其中一份置于密闭遮光的真空干燥器中, 用无酒精的氯仿熏蒸24 小时, 另一份不进行熏蒸; 通风, 待熏蒸组氯仿散尽, 两份土壤样品均用 25mL 浓度为 0.5M 的K2SO4 溶液浸提。用总有机碳分析仪(Multi N/C® 3100, Analytik Jena AG, Jena, 德国)测定两份土壤浸提液中碳含量, 二者的差值为土壤微生物体内的碳含量, 以 0.45 作为修正系数, 对微生物量碳含量进行校正[30]。用元素分析仪(Vario EL III, Ele- mentar, 德国)测定土壤总碳和总氮含量, 用温湿度传感器(POGO mini, Stevens, 美国)在野外测定土壤温度, 用烘干称重法进行测定土壤含水量, 用粒度分析仪(Mastersizer 2000; Malvern, Wor-cestershire, 英国)测定土壤粒径, 用 pH 计(Model PHS-2; 上海仪电科学仪器股份有限公司, 中国)测定土壤 pH值。两个土层的土壤理化性质见表 2。
图1 样点位置
Fig. 1 Locations of the sampling sites
表1 样点信息
Table 1 Information of sampling sites
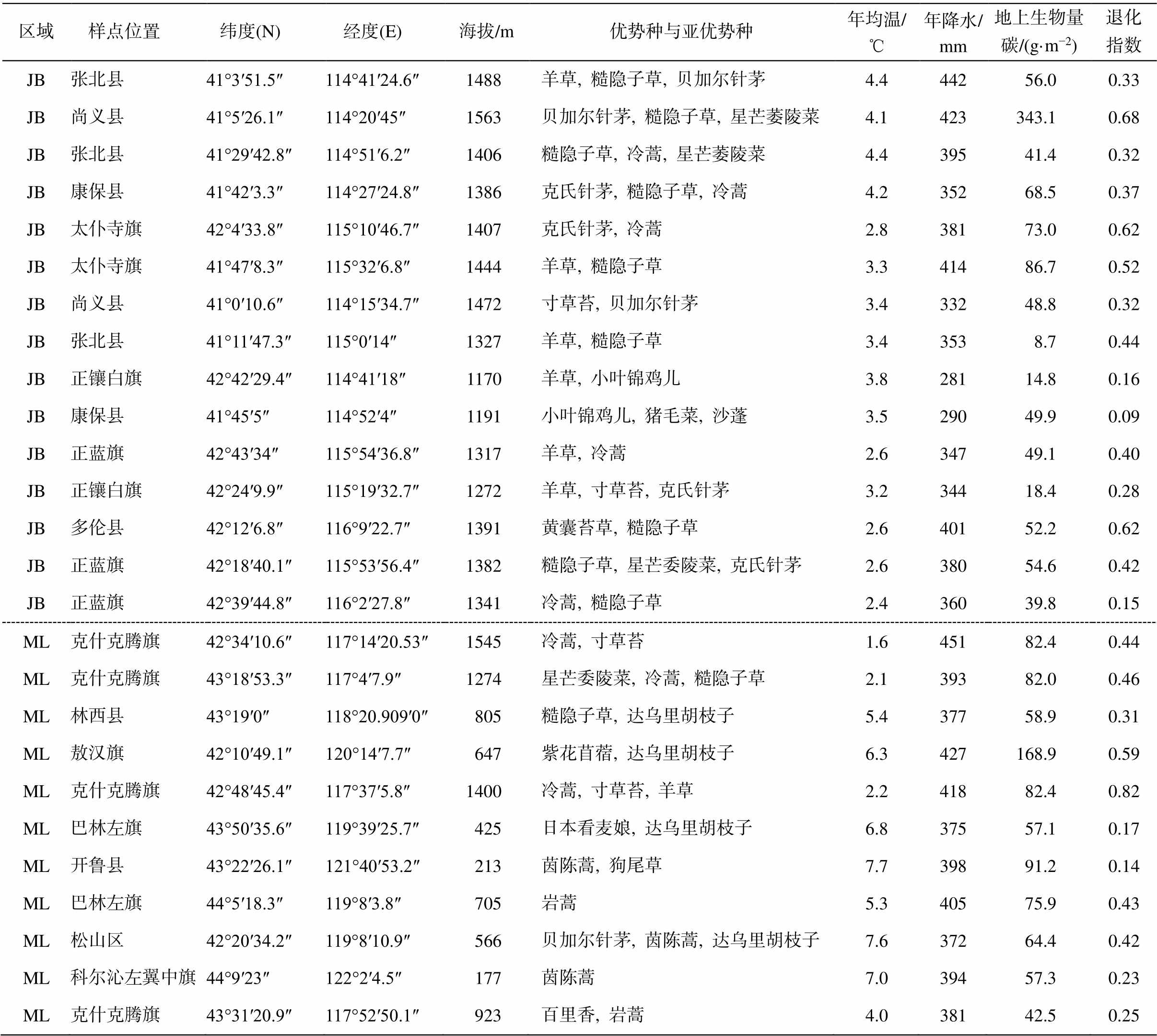
区域 样点位置 纬度(N) 经度(E)海拔/m优势种与亚优势种年均温/℃年降水/ mm地上生物量碳/(g·m−2)退化指数JB张北县41°3′51.5″114°41′24.6″1488羊草, 糙隐子草, 贝加尔针茅4.444256.00.33JB尚义县41°5′26.1″114°20′45″1563贝加尔针茅, 糙隐子草, 星芒萎陵菜4.1423343.10.68JB张北县41°29′42.8″114°51′6.2″1406糙隐子草, 冷蒿, 星芒萎陵菜4.439541.40.32JB康保县41°42′3.3″114°27′24.8″1386克氏针茅, 糙隐子草, 冷蒿4.235268.50.37JB太仆寺旗42°4′33.8″115°10′46.7″1407克氏针茅, 冷蒿2.838173.00.62JB太仆寺旗41°47′8.3″115°32′6.8″1444羊草, 糙隐子草3.341486.70.52JB尚义县41°0′10.6″114°15′34.7″1472寸草苔, 贝加尔针茅3.433248.80.32JB张北县41°11′47.3″115°0′14″1327羊草, 糙隐子草3.43538.70.44JB正镶白旗42°42′29.4″114°41′18″1170羊草, 小叶锦鸡儿3.828114.80.16JB康保县41°45′5″114°52′4″1191小叶锦鸡儿, 猪毛菜, 沙蓬3.529049.90.09JB正蓝旗42°43′34″115°54′36.8″1317羊草, 冷蒿2.634749.10.40JB正镶白旗42°24′9.9″115°19′32.7″1272羊草, 寸草苔, 克氏针茅3.234418.40.28JB多伦县42°12′6.8″116°9′22.7″1391黄囊苔草, 糙隐子草2.640152.20.62JB正蓝旗42°18′40.1″115°53′56.4″1382糙隐子草, 星芒委陵菜, 克氏针茅2.638054.60.42JB正蓝旗42°39′44.8″116°2′27.8″1341冷蒿, 糙隐子草2.436039.80.15ML克什克腾旗42°34′10.6″117°14′20.53″1545冷蒿, 寸草苔1.645182.40.44ML克什克腾旗43°18′53.3″117°4′7.9″1274星芒委陵菜, 冷蒿, 糙隐子草2.139382.00.46ML林西县43°19′0″118°20.909′0″805糙隐子草, 达乌里胡枝子5.437758.90.31ML敖汉旗42°10′49.1″120°14′7.7″647紫花苜蓿, 达乌里胡枝子6.3427168.90.59ML克什克腾旗42°48′45.4″117°37′5.8″1400冷蒿, 寸草苔, 羊草2.241882.40.82ML巴林左旗43°50′35.6″119°39′25.7″425日本看麦娘, 达乌里胡枝子6.837557.10.17ML开鲁县43°22′26.1″121°40′53.2″213茵陈蒿, 狗尾草7.739891.20.14ML巴林左旗44°5′18.3″119°8′3.8″705岩蒿5.340575.90.43ML松山区42°20′34.2″119°8′10.9″566贝加尔针茅, 茵陈蒿, 达乌里胡枝子7.637264.40.42ML科尔沁左翼中旗44°9′23″122°2′4.5″177茵陈蒿7.039457.30.23ML克什克腾旗43°31′20.9″117°52′50.1″923百里香, 岩蒿4.038142.50.25
续表

区域 样点位置 纬度(N) 经度(E)海拔/m优势种与亚优势种年均温/℃年降水/ mm地上生物量碳/(g·m−2)退化指数ML扎鲁特旗44°31′23.9″120°49′0.9″287茵陈蒿, 糙隐子草, 达乌里胡枝子7.140376.90.79ML阜新县42°25′34.7″121°21′13.4″321糙隐子草, 日本看麦娘7.342235.80.18ML巴林右旗43°38′59.1″118°35′23″906糙隐子草, 达乌里胡枝子, 狼毒4.640560.40.35ML奈曼旗43°24′57.8″120°47′19.8″288蒺藜, 茵陈蒿, 狗尾草7.638036.00.11YS察哈尔右翼前旗40°48′45.7″113°29′30.4″1374克氏针茅, 扁蓿豆5.538767.90.27YS卓资县40°59′32.6″112°35′5.3″1563高裂委陵菜, 冷蒿, 羊草4.340130.10.59YS四子王旗41°42.605′″111°46.287′″1466寸草苔, 轴黎, 克氏针茅4.629238.40.28YS托克托40°10′43.7″111°13′56.4″1065本氏针茅, 兴安胡枝子8.641782.00.36YS达拉特旗40°14′22″110°37′22.7″1013尖头叶藜, 肉叶雾冰藜, 狗尾草9.039257.30.13YS青山区40°41′21.4″109°56′37.4″1112本氏针茅8.232971.90.48YS达拉特旗40°10′2.8″109°23′52.4″1259克氏针茅, 牛心朴子8.533737.60.36YS东胜区39°48′26.8″109°37′17.6″1480克氏针茅, 狗牙花, 兴安胡枝子7.140592.10.28YS神木市38°47′33.6″110°13′37.5″1261野谷草, 沙蒿, 茵陈蒿8.8514186.10.27YS府谷县38°57′41.7″110°50′59.3″1146地梢瓜, 硬质早熟禾, 兴安胡枝子9.349795.30.29YS清水河县39°44′28.6″111°44′28.6″1248大针茅, 本氏针茅, 斜茎黄耆, 兴安胡枝子8.0460109.60.46YS平鲁区39°46′12.1″112°6′41.3″1578硬质早熟禾, 直立地蔷薇, 大针茅5.9487168.20.39YS和林格尔县40°15′32″112°12′18.5″1274硬质早熟禾, 扁蓿豆, 草木犀状黄芪6.643466.80.34YS右玉县40°0′20.9″112°35′16.8″1470星毛委陵菜, 冷蒿, 克氏针茅6.3456118.50.48NS乌审旗38°48′11.9″109°7′2.3″1297羊草, 因陈蒿, 芦苇8.441394.90.29NS鄂托克旗38°47′31.4″108°25′43.1″1387兴安胡枝子, 狗尾草, 肉叶雾冰藜7.9359109.30.15NS鄂托克前旗38°16′26.4″107°30′40.5″1320糙隐子草, 茵陈蒿, 阿尔泰狗尾花9.031962.10.23NS鄂托克前旗38°21′53.8″106°41′23.9″1303野谷草, 本氏针茅, 茵陈蒿9.525733.40.24NS灵武市37°43′58.6″106°28′40″1266油蒿, 牛心朴子, 刺旋花10.126699.20.24NS同心县36°59′58.5″106°24′51.8″1772栉叶蒿, 茵陈蒿, 赖草, 羊草7.7386110.50.48NS海原县36°55′2.2″105°16′15.2″1729茵陈蒿, 兴安胡枝子, 节毛飞镰8.428742.30.34NS红寺堡区37°25′10.4″105°51′57.5″1313茵陈蒿, 葡枝骆驼莲, 刺旋花10.324640.90.30NS定边县37°42′43.6″107°30′8.1″1310茵陈蒿, 兴安胡枝子, 牛心朴子9.336397.30.22NS定边县37°28′59.5″108°15′57.4″1467硬质早熟禾, 克氏针茅, 糙隐子草7.949980.60.34NS乌审旗37°59′1.6″108°44′3.4″1188茵陈蒿, 沙韭, 硬质早熟禾, 砂珍棘豆9.4426214.50.22NS乌审旗38°31′49.7″108°49′48.8″1301蓝刺头, 砂珍棘豆, 狗尾巴草8.7408196.10.12NS榆阳区38°15′59.3″109°38′29.8″1152草木犀状黄芪, 沙蒿, 茵陈蒿9.5475151.40.25
用文献[18,31]中建立的包含植物和土壤特征的综合性草地退化指数(grassland degradation index, GDI), 对退化程度进行量化, GDI 指数越低, 表征草地退化程度越强。其中, 植物特征包括一年生植物、退化指示种和顶极种的盖度[32], 土壤特征包括土壤总碳、总氮和砂粒含量, 计算公式如下:
表2 表层与次表层土壤的理化性质(平均值±标准误差)
Table 2 Soil physiochemical properties of surface and subsurface layers (mean±SE)

土层土壤全碳/(mmol·kg −1)土壤全氮/(mmol·kg −1)黏粒含量/%土壤温度/(℃)土壤含水量/%pH0~10 cm1122.32±89.7074.63±7.907.24±0.2930.32±0.444.54±0.058.07±0.1010~20 cm1033.28±86.8849.86±5.907.91±0.3230.48±0.455.81±0.048.14±0.09
GDI=(P1×1/3+P2×2/3+P3)×1/3+
(STC×1/2+STN×1/2)×1/3+(1/Sand)×1/3, (1)
其中, P1, P2 和 P3 分别表示一年生植物、退化指示种和顶极种的盖度, STC 和 STN 分别表示土壤总碳和总氮含量, Sand 表示土壤砂粒含量。
次表层土壤微生物量碳相对于表层土壤的变化率(change rate, CR)计算公式如下:
CR = (MBC10~20cm − MBC0~10cm) / MBC0~10cm, (2)
其中, MBC10~20cm 和 MBC0~10cm 分别表示次表层和表层土壤微生物量碳含量。CR 越接近 0, 表示次表层与表层微生物量碳的差异越小; CR 为负值表示次表层土壤微生物量碳含量高于表层土壤。
采用线性回归方法, 分析微生物量碳的空间格局及其变化率与环境因素的关系。采用 Duncan’s多重分析方法, 比较不同区域微生物量碳差异的显著性。采用配对 t 检验方法, 分析表层与次表层土壤微生物量碳的差异。用 Pearson 相关方法, 分析微生物量碳与环境因素的关系。使用 randomForest包进行随机森林分析, 对环境因素的重要性指数进行排序。用 SPSS24.0 和 R3.6 软件进行数据分析, 使用 GraphpadPrism8 作图。显著性检验的标准为P<0.05。
表层与次表层土壤微生物量碳均随着纬度的增加而显著增加(图 2(a)), 且变化幅度相近。随着经度和海拔增长, 表层与次表层微生物量碳仅呈现增加趋势, 检验结果均不显著(P>0.05, 图 2(b)和(c))。
两个土层的分析结果均显示, 京北区域的土壤微生物量碳含量显著地高于宁陕区域, 蒙辽区域与阴山区域无显著差异(图 3(a))。总体而言, 农牧交错带草地表层与次表层土壤微生物量碳的空间格局具有一致性。
本文发现农牧交错带草地表层与次表层土壤微生物量碳随纬度增加而增加, 与 Xu 等[6]的研究结果一致。随着纬度增加, 年均温逐渐降低, 导致有机质分解速率减小, 有利于土壤中碳和氮的累积[33]。区域[16]和全球[34-35]的研究均发现, 土壤碳含量与微生物量碳正相关。随着纬度的变化, 年均温等环境因素发生显著的改变[36], 从而对土壤微生物产生影响, 相比之下, 农牧交错带微生物量碳随经度的变化并不显著。本文研究区域农牧交错带草地表层与次表层土壤微生物量碳随海拔的变化均不显著, 原因可能是北方农牧交错带位于温带, 而非热带-亚热带。He 等[10]的全球整合分析结果表明, 仅热带和亚热带区域土壤微生物量碳随海拔升高而显著升高, 温带和寒带无显著变化。农牧交错带草地表层和次表层土壤微生物量碳随经纬度和海拔的变化呈现一致的规律。
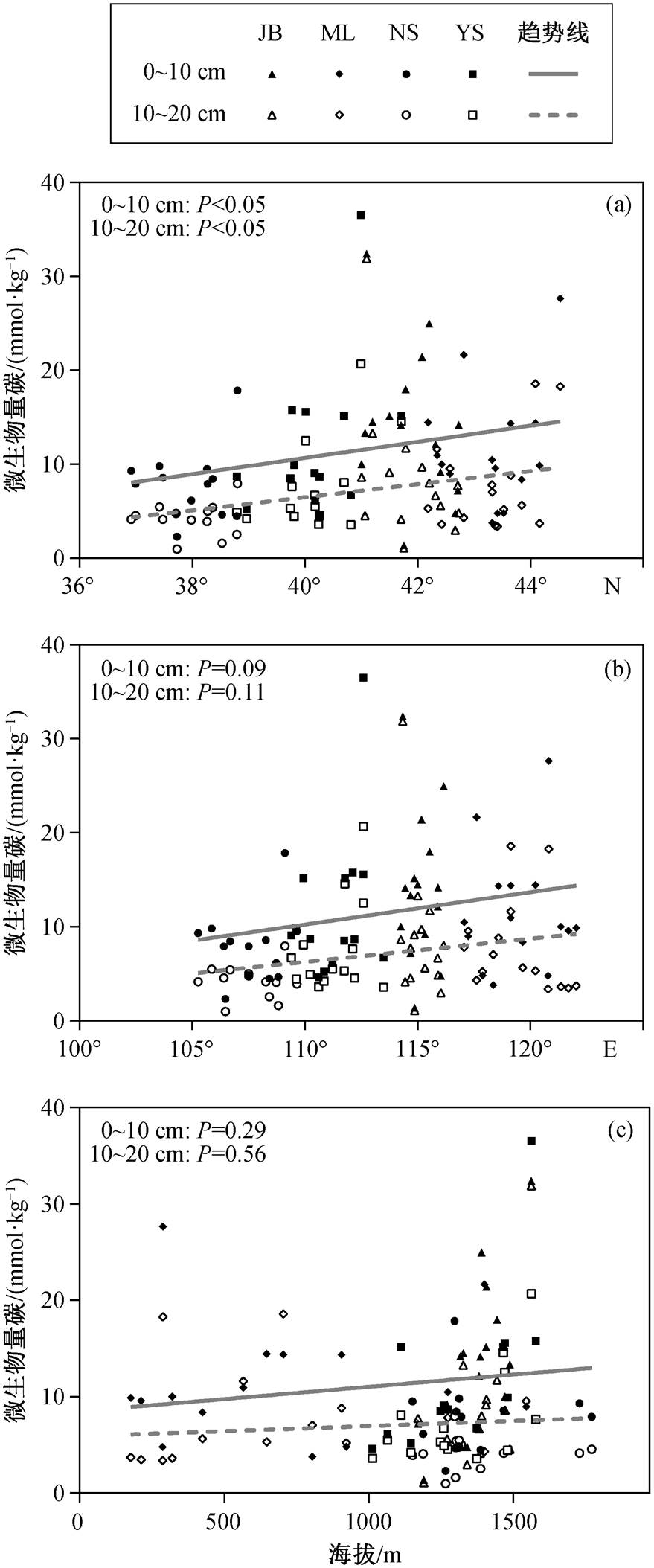
图2 表层与次表层土壤微生物量碳的空间分布
Fig. 2 Spatial pattern of microbial biomass carbon in surface and subsurface soils
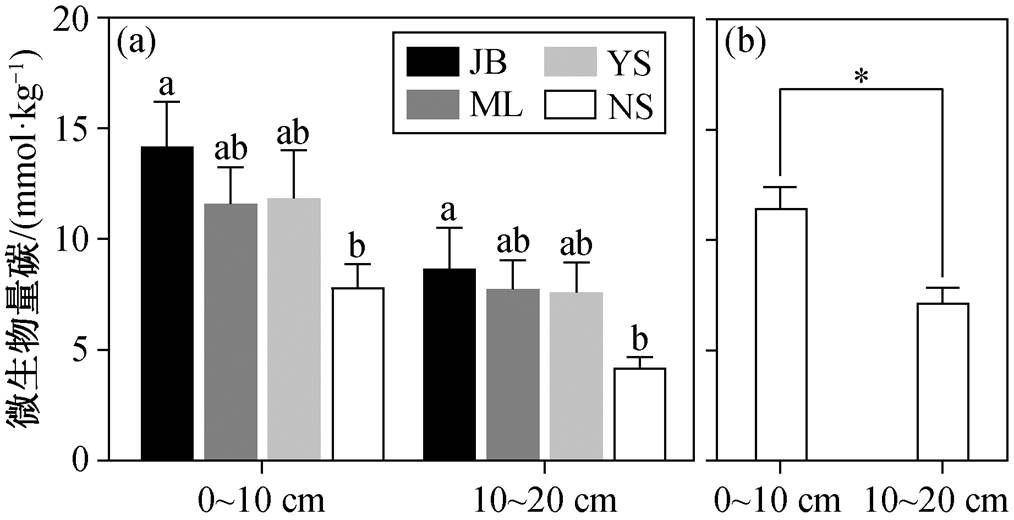
不同小写字母表示同一深度不同区域之间差异的显著性(Duncan’s多重比较), 误差棒代表标准误差, *表示P<0.05
图3 不同区域(a)及所有样点(b)表层与次表层微生物量碳
Fig. 3 Surface and subsurface layer microbial biomass carbon of different areas (a) and all sample points (b)
随着草地退化程度加剧, 表层与次表层土壤微生物量碳均显著降低(图 4(a)), 表层土壤微生物量碳的变化幅度更大(表层和次表层土壤微生物量碳的变化斜率分别为 33.61 和 19.33)。虽然区域表层土壤微生物量碳均值整体上高于次表层土壤(图 3 (b)), 但表层与次表层土壤微生物量碳的差异并不恒定, 19.3%的样点次表层土壤微生物量碳含量高于表层土壤。草地退化指数和微生物量碳变化率的回归分析结果显示, 随着草地退化程度的加剧, 次表层与表层土壤微生物量碳的差异呈现降低的趋势, 但降低幅度不显著(图 4(b))。
回归分析结果显示, 土壤微生物量碳变化率与土壤含水量变化率、土壤温度变化率、黏粒含量变化率、土壤总碳变化率和土壤总氮变化率均无显著关系(图 5(a)~(e)), 而与土壤 pH 值变化率显著负相关(P<0.05, 图 5(f))。进一步分析发现, 土壤 pH 值变化率与草地退化程度显著相关(图 6), 随着退化程度加剧, 表层与次表层的土壤 pH 值差异减小。
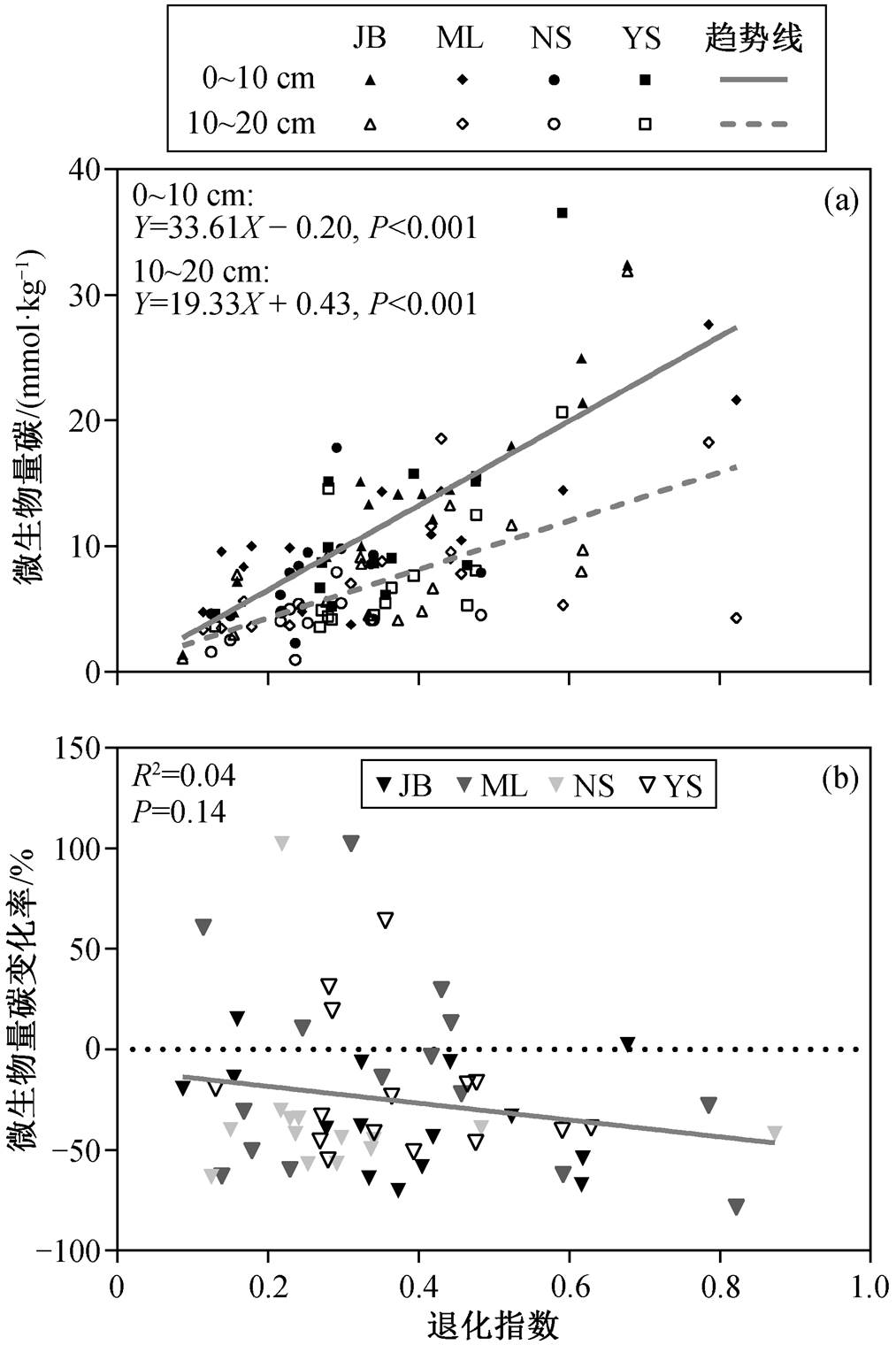
图4 草地退化指数与微生物量碳(a)及微生物量碳变化率(b)的关系
Fig. 4 Relationship between GDI and MBC (a) and the change rate of MBC (b)
本文研究结果表明, 表层和次表层土壤微生物量碳均随草地退化程度加剧而显著降低。一方面, 随着草地退化程度加剧, 植被覆盖度与生物量碳显著降低[37], 导致植物碳输入减少, 表层土壤直接裸露在空气中, 土壤水分加速流失[38], 不利于土壤微生物生长[23]。另一方面, 草地退化还导致土壤理化性质改变, 如土壤沙化和养分流失[37]。此外, 放牧活动中的牲畜踩踏使土壤孔隙度降低[39], 通气性变差[40], 导致土壤微生物量碳含量降低[11]。本文还发现, 随草地退化的加剧, 次表层土壤微生物量碳变化幅度小于表层土壤。彭晓茜等[17]对内蒙古温带草原的研究结果显示, 环境因素对微生物量碳的影响随土壤深度的增加而减弱。表层土壤受到环境的直接干扰[41], 对次表层土壤起到缓冲作用, 因此次表层土壤环境相对稳定[42], 受草地退化的影响弱于表层土壤。
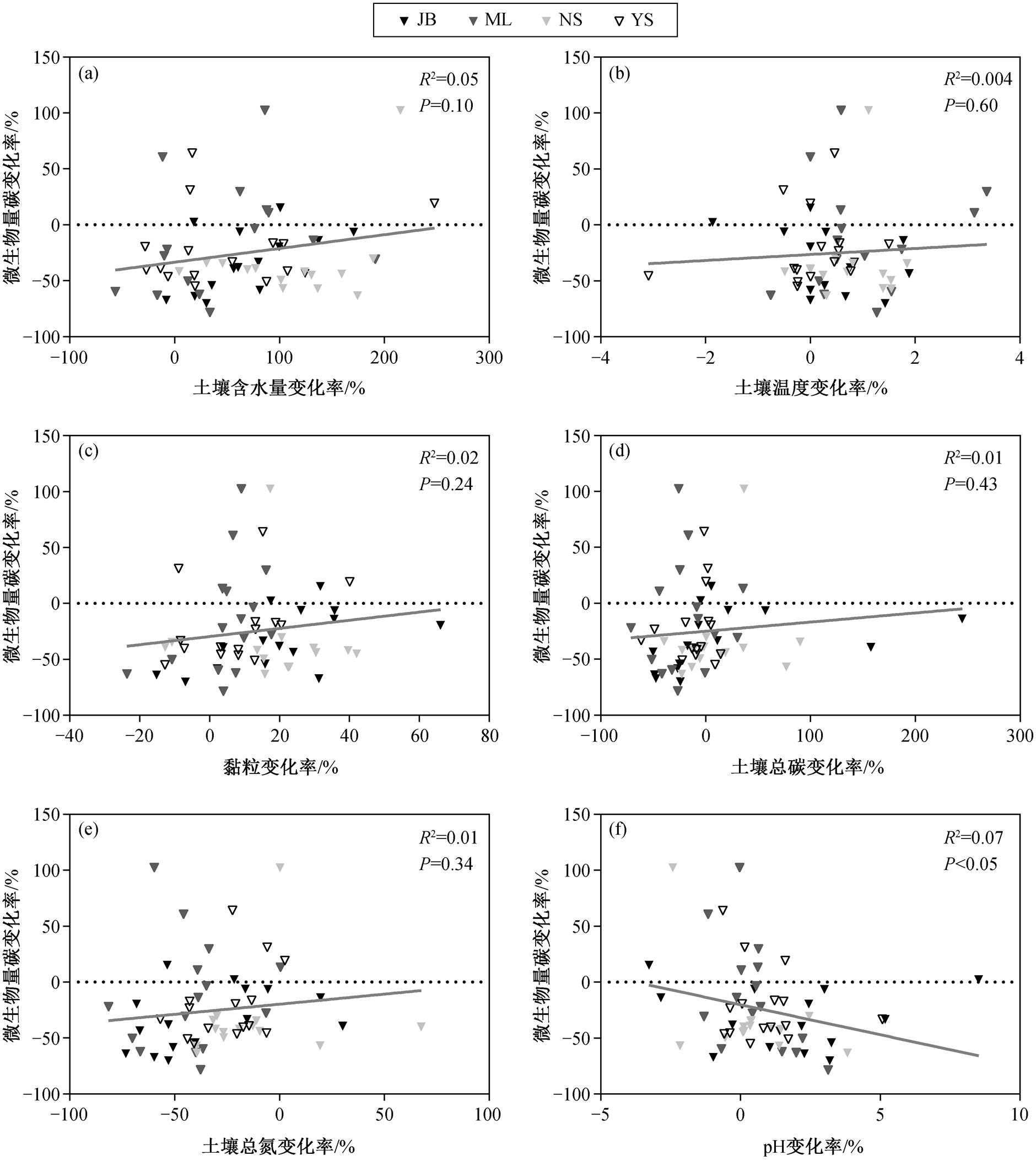
图5 微生物量碳变化率与环境因素变化率的关系
Fig. 5 Relationship between relative change rate of MBC and change rate of environmental factors
微生物量碳变化率分析结果显示, 随着草地退化程度增加, 次表层与表层土壤微生物量碳的差异呈现降低的趋势, 与贺凤鹏等[23]对内蒙古不同退化程度草地的研究结果一致。本文发现土壤微生物量碳变化率与土壤 pH 变化率显著负相关, 表明表层与次表层土壤 pH 值的差异越小, 微生物量碳的差异也越小。土壤 pH 值是调控微生物群落组成及活性的重要非生物因素之一, 对微生物量碳具有较大的影响[41-44]。随着土壤 pH 增加, 真菌与细菌的比例升高[45], 真菌具有比细菌更多的碳需求[46], 从而影响微生物量碳含量[47-48]。Sinsabaugh 等[49]对全球土壤微生物碳利用效率的分析发现, 当土壤 pH>7时, 微生物的碳利用效率随 pH 的增加而上升, 意味着微生物将更多的碳用于自身生长而非呼吸代谢, 因此能够增加微生物量碳。回归分析结果显示, 随着草地退化指数降低, 表层与次表层土壤 pH 的差异越来越小, 表明草地退化导致两个土层之间 pH的差异减小, 进而使两个土层之间微生物量碳的差异减小。因此, 土壤 pH 的差异是导致区域尺度表层与次表层微生物量碳差异的主要原因。
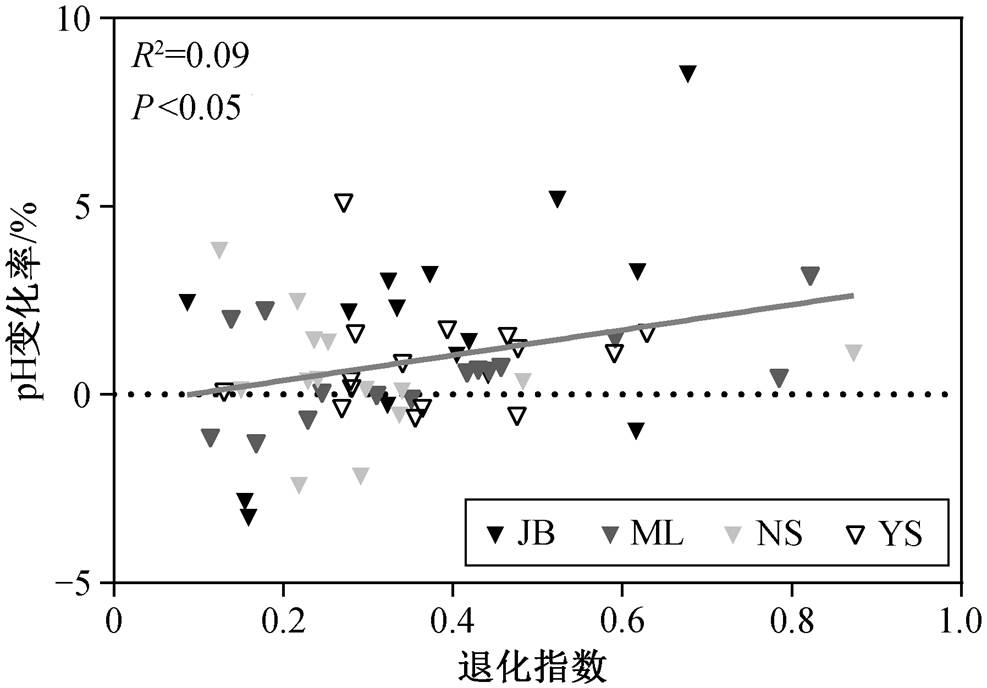
图6 土壤pH值变化率与草地退化指数的关系
Fig. 6 Relationship between change rate of soil pH and GDI
本文发现, 在区域尺度上, 退化程度较高(退化指数<0.5)的区域共有 10 个样点的次表层土壤微生物量碳高于表层土壤。一方面, 随着草地退化程度加剧, 表层土壤沙化程度加重, 保水能力降低, 不利于维持微生物生长[23]; 另一方面, 由于退化程度高的区域部分样点次表层土壤的 pH 高于表层土壤, 而高 pH 值有利于提高微生物碳利用效率[49], 因此部分样点次表层土壤含有更多的微生物量碳。
皮尔森相关性分析结果(表 3)显示, 气候、植被和土壤因素均对微生物量碳有显著影响。表层和次表层土壤微生物量碳均与土壤总碳、土壤总氮、黏粒含量和土壤含水量显著正相关, 与年均温显著负相关。
用随机森林算法对各环境因素的重要性进行排序, 结果显示土壤因素对微生物量碳的影响最大(图 7)。对于表层土壤微生物量碳, 土壤总碳是最重要的影响因素, 其次是土壤总氮和土壤含水量(图 7(a)); 对于次表层土壤微生物量碳, 土壤总氮是最主要的影响因素, 其次是土壤含水量, 黏粒对土壤微生物量碳的贡献与土壤总碳相当(图 7(b))。
本文研究结果显示, 土壤、气候和植被因素均对微生物量碳产生影响。气候能够通过影响植被生长和土壤含水量等, 间接地影响微生物量碳[13]。植被因素也与土壤微生物生长显著相关, 例如较高的根系生物量伴随着较高的根系分泌物, 有利于微生物的生长[6,17]。进一步的分析表明, 土壤因素对微生物量碳的影响大于气候和植被因素, 这一结果与Hu 等[16]对干旱和半干旱区草地的研究结果一致。Zhao 等[13]通过对高寒草地与温带草地的整合研究, 发现气候因素的作用显著大于土壤因素, 原因可能是高寒草地年均温低[50], 低温成为微生物生长的限制因素[51]。此外, 气候和植物因素可能通过影响土壤因素间接地作用于微生物量碳[16], 因此造成土壤因素表现出最强的直接影响。作为微生物代谢能量和营养的基本来源[52], 土壤碳和氮与微生物量碳显著正相关[10]。土壤含水量直接影响微生物的生理和生长[48], 农牧交错带降水有限, 微生物生长受土壤含水量限制, 因此与土壤含水量显著正相关。黏粒能够保护微生物免受捕食[53], 同时提高土壤的持水能力[54], 因此质地良好的土壤具有更高的微生物量碳。
本文还发现, 表层土壤微生物量碳的主要环境驱动力为土壤总碳。以往的研究表明, 土壤碳含量可以作为微生物量碳空间格局的良好预测指标[6,10]。但是, 与表层土壤不同, 次表层土壤微生物量碳的主要驱动力为土壤总氮, 原因可能是次表层土壤的养分限制强于能量限制。Liang 等[55]通过温带草地土壤养分添加培养实验发现, 相对于表层土壤, 深层土壤微生物存在更强的氮限制。微生物代谢效率随着氮可利用性的降低而降低[56], 意味着微生物将在氮限制的情况下, 将更多的碳分配给呼吸而非生长, 进而影响微生物量碳含量。因此, 对于次表层土壤的微生物量碳的预测, 应将土壤总氮含量也作为关键驱动因子纳入考虑。
表3 微生物量碳与环境因素的皮尔森相关性分析结果
Table 3 Results of Pearson correlation coefficients between microbial biomass carbon and environmental factors

土层气候因素土壤因素植物因素MATMAPSTCSTNCLAYSTSMpHAB0~10 cm−0.326*0.2420.757**0.704**0.491**−0.1390.729**−0.1930.20210~20 cm−0.306*0.1750.647**0.741**0.512**−0.1260.682**−0.1590.270*
说明: MAT表示年均温, MAP表示年降水, STC表示土壤全碳含量, STN表示土壤全氮含量, CLAY表示黏粒含量, ST表示土壤温度, SM表示土壤含水量, AB表示地上生物量碳, 下同; *表示P<0.05, **表示P<0.01。
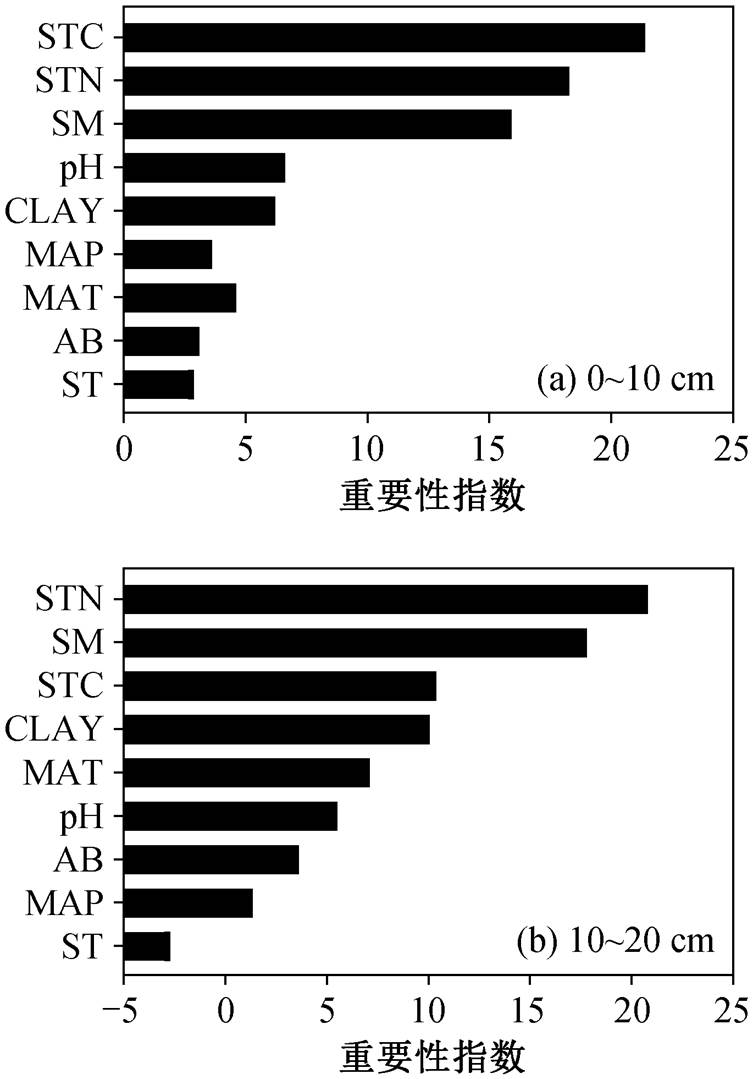
图7 表层(a)与次表层(b)微生物量碳与环境因素的随机森林分析结果
Fig. 7 Results of random forest analysis of microbial biomass carbon and environmental factors in surface (a) and subsurface (b) layer
本文利用我国北方农牧交错带 4 个区域 57 个草地样点的 456 个土壤样品, 研究农牧交错带草地表层和次表层土壤微生物量碳的分布格局及环境驱动力, 得到如下结论。
1)表层与次表层土壤微生物量碳具有一致的空间格局, 均随纬度的增加而增加, 随草地退化程度的加重而降低。
2)表层与次表层土壤 pH 的差异随草地退化程度的加剧而缩小, 进而使两个土层的微生物量碳差异减小。
3)土壤因素是微生物量碳空间分布的主要驱动力。
4)表层土壤微生物量碳受土壤总碳的影响最大, 次表层土壤微生物量碳主要受土壤总氮影响。
本文研究结果有利于深入认识区域尺度草地生态系统的生物地球化学循环, 为退化草地的生态功能恢复提供科学依据。
参考文献
[1] Zechmeister-Boltenstern S, Keiblinger K M, Moosha-mmer M, et al. The application of ecological stoichio-metry to plant-microbial-soil organic matter transfor-mations. Ecological Monographs, 2015, 85(2): 133–155
[2] Trivedi P, Anderson I C, Singh B K. Microbial modu-lators of soil carbon storage: integrating genomic and metabolic knowledge for global prediction. Trends in Microbiology, 2013, 21(12): 641–651
[3] Spohn M, Kuzyakov Y. Phosphorus mineralization can be driven by microbial need for carbon. Soil Bio-logy and Biochemistry, 2013, 61: 69–75
[4] Li Zhaolei, Tian Dashuan, Wang Bingxue, et al. Mic-robes drive global soil nitrogen mineralization and availability. Global Change Biology, 2019, 25: 1078–1088
[5] Spohn M. Element cycling as driven by stoichiometric homeostasis of soil microorganisms. Basic and Applied Ecology, 2016, 17(6): 471–478
[6] Xu X F, Thornton P E, Post W M. A global analysis of soil microbial biomass carbon, nitrogen and phos-phorus in terrestrial ecosystems. Global Ecology and Biogeography, 2013, 22(6): 737–749
[7] Joergensen R G, Wichern F. Alive and kicking: why dormant soil microorganisms matter. Soil Biology and Biochemistry, 2018, 116: 419–430
[8] Miltner A, Bombach P, Schmidt-Brücken B, et al. SOM genesis: microbial biomass as a significant source. Biogeochemistry, 2012, 111(1/2/3): 41–55
[9] Kallenbach C M, Frey S D, Grandy A S. Direct evi-dence for microbial-derived soil organic matter forma-tion and its ecophysiological controls. Nature Com-munications, 2016, 7: 1–10
[10] He Xianjin, Hou Enqing, Veen G F, et al. Soil micro-bial biomass increases along elevational gra-dients in the tropics and subtropics but not elsewhere. Global Ecology and Biogeography, 2019, 29: 345–354
[11] Singh J S, Gupta V K. Soil microbial biomass: a key soil driver in management of ecosystem functioning. Science of the Total Environment, 2018, 634: 497–500
[12] Li Xiangzhen, Chen Zuozhong. Soil microbial bio-mass C and N along a climatic transect in the Mongo-lian steppe. Biology and Fertility of Soils, 2004, 39 (5): 344–351
[13] Zhao Huan, Sun Jian, Xu Xingliang, et al. Stoichio-metry of soil microbial biomass carbon and microbial biomass nitrogen in China’s temperate and alpine grasslands. European Journal of Soil Biology, 2017, 83: 1–8
[14] Šnajdr J, Valášková V, Merhautová V, et al. Spatial variability of enzyme activities and microbial biomass in the upper layers of Quercus petraea forest soil. Soil Biology and Biochemistry, 2008, 40(9): 2068–2075
[15] Peng Xiaoqian, Wang Wei. Stoichiometry of soil ex-tracellular enzyme activity along a climatic transect in temperate grasslands of northern China. Soil Biology and Biochemistry, 2016, 98: 74–84
[16] Hu Yajun, Xiang Dan, Veresoglou S D, et al. Soil organic carbon and soil structure are driving micro-bial abundance and community composition across the arid and semi-arid grasslands in northern China. Soil Biology and Biochemistry, 2014, 77: 51–57
[17] 彭晓茜, 王娓. 内蒙古温带草原土壤微生物生物量碳的空间分布及驱动因素. 微生物学通报, 2016, 43(9): 1918–1930
[18] Zeng Wenjing, Chen Jinbin, Liu Hongyan, et al. Soil respiration and its autotrophic and heterotrophic com-ponents in response to nitrogen addition among dif-ferent degraded temperate grasslands. Soil Biology and Biochemistry, 2018, 124(2): 255–265
[19] Akiyama T, Kawamura K. Grassland degradation in China: methods of monitoring, management and res-toration. Grassland Science, 2007, 53(1): 1–17
[20] Li Yuefen, Li Qingyu, Guo Dongyan, et al. Ecological stoichiometry homeostasis of Leymus chinensis in de-graded grassland in western Jilin Province, NE China. Ecological Engineering, 2016, 90: 387–391
[21] Zhou Zhenghu, Wang Chuankuan, Luo Yiqi. Effects of forest degradation on microbial communities and soil carbon cycling: a global meta-analysis. Global Ecology and Biogeography, 2018, 27(1): 110–124
[22] 杨倩, 王娓, 曾辉. 氮添加对内蒙古退化草地植物群落多样性和生物量的影响. 植物生态学报, 2018, 42(4): 430–441
[23] 贺凤鹏, 曾文静, 王曌迪, 等. 温带草原退化对土壤剖面微生物学特征的影响. 微生物学通报, 2016, 43(3): 702–711
[24] Huang Ding, Wang Kun, Wu Wenliang. Problems and strategies for sustainable development of farming and animal husbandry in the Agro-Pastoral Transition Zone in Northern China (APTZNC). International Journal of Sustainable Development and World Ecology, 2007, 14(4): 391‒399
[25] 李旭亮, 杨礼箫, 田伟, 等. 中国北方农牧交错带土地利用/覆盖变化研究综述. 应用生态学报, 2018, 29(10): 331–339
[26] 苏伟, 陈云浩, 武永峰, 等. 生态安全条件下的土地利用格局优化模拟研究——以中国北方农牧交错带为例. 自然科学进展, 2006, 16(2): 207–214
[27] 李栋梁, 吕兰芝. 中国农牧交错带的气候特征与演变. 中国沙漠, 2002, 22(5): 76–81
[28] Liu Xu, Liu Hongyan, Qiu Shuang, et al. An improved estimation of regional fractional woody/herbaceous cover using combined satellite data and high-quality training samples. Remote Sensing, 2017, 9(1): 1–13
[29] Vance E D, Brookes P C, Jenkinson D S. An extra-ction method for measuring soil microbial biomass C. Soil Biology and Biochemistry, 1987, 19(6): 703–707
[30] Jenkinson D S, Powlson D S. The effects of biocidal treatments on metabolism in soil — V: a method for measuring soil biomass. Soil Biology and Bioche-mistry, 1976, 8(3): 209–213
[31] Dong Chengcheng, Wang Wei, Liu Hongyan, et al. Temperate grassland shifted from nitrogen to phosp-horus limitation induced by degradation and nitrogen deposition: evidence from soil extracellular enzyme stoichiometry. Ecological Indicators, 2019, 101(2): 453–464
[32] Liu Hongyan, Yin Yi, Tian Yuhong, et al. Climatic and anthropogenic controls of topsoil features in the semi-arid East Asian steppe. Geophysical Research Letters, 2008, 35(4): 1–5
[33] Garten C T, Hanson P J. Measured forest soil C stocks and estimated turnover times along an elevation gra-dient. Geoderma, 2006, 136(1): 342–352
[34] Cleveland C C, Liptzin D. C:N:P stoichiometry in soil: is there a “Redfield ratio” for the microbial biomass?. Biogeochemistry, 2007, 85(3): 235–252
[35] Fierer N, Strickland M S, Liptzin D, et al. Global patterns in belowground communities. Ecology Let-ters, 2009, 12(11): 1238–1249
[36] Chen Yongliang, Chen Leiyi, Peng Yunfeng, et al. Linking microbial C:N:P stoichiometry to microbial community and abiotic factors along a 3500-km grassland transect on the Tibetan Plateau. Global Ecology and Biogeography, 2016, 25(12): 1416–1427
[37] 郝爱华, 薛娴, 彭飞, 等. 青藏高原典型草地植被退化与土壤退化研究. 生态学报, 2018, 40(3): 964–975
[38] Xue Xian, You Quangang, Peng Fei, et al. Experi-mental warming aggravates degradation-induced top-soil drought in alpine meadows of the Qinghai-Tibetan Plateau. Land Degradation & Development, 2017, 28(8): 2343–2353
[39] 王蕙, 王辉, 黄蓉, 等. 不同封育管理对沙质草地土壤与植被特征的影响. 草业学报, 2012, 21(6): 15–22
[40] 王群, 尹飞, 郝四平, 等. 下层土壤容重对玉米根际土壤微生物数量及微生物量碳、氮的影响. 生态学报, 2009, 29(6): 3096–3104
[41] Yao Xiaodong, Zhang Naili, Zeng Hui, et al. Effects of soil depth and plant–soil interaction on microbial community in temperate grasslands of northern China. Science of the Total Environment, 2018, 630: 96–102
[42] Yang Yuanhe, Fang Jinyun, Guo Dali, et al. Vertical patterns of soil carbon, nitrogen and carbon: nitrogen stoichiometry in Tibetan grasslands. Biogeosciences Discussions, 2010, 7(1): 1–24
[43] Rousk J, Brookes P C, Bååth E. The microbial PLFA composition as affected by pH in an arable soil. Soil Biology and Biochemistry, 2010, 42(3): 516–520
[44] Li Pin, Yang Yuanhe, Han Wenxuan, et al. Global patterns of soil microbial nitrogen and phosphorus stoichiometry in forest ecosystems. Global Ecology and Biogeography, 2014, 23(9): 979–987
[45] Bååth E, Anderson T H. Comparison of soil fungal/ bacterial ratios in a pH gradient using physiological and PLFA-based techniques. Soil Biology and Bio-chemistry, 2003, 35(7): 955–963
[46] Keiblinger K M, Hall E K, Wanek W, et al. The effect of resource quantity and resource stoichiometry on microbial carbon-use-efficiency. FEMS Microbiology Ecology, 2010, 73(3): 430–440
[47] Tian Ye, Haibara K, Toda H, et al. Microbial biomass and activity along a natural pH gradient in forest soils in a karst region of the upper Yangtze River, China. Journal of Forest Research, 2008, 13(4): 205–214
[48] Manzoni S, Taylor P, Richter A, et al. Environmental and stoichiometric controls on microbial carbon-use efficiency in soils. New Phytologist, 2012, 196(1): 79–91
[49] Sinsabaugh R L, Turner B L, Talbot J M, et al. Stoichiometry of microbial carbon use efficiency in soils. Ecological Monographs, 2016, 86(2): 172–189
[50] Sun Jin, Cheng Genwei, Li Weipeng. Meta-analysis of relationships between environmental factors and aboveground biomass in the alpine grassland on the Tibetan Plateau. Biogeosciences, 2013, 10(3): 1707–1715
[51] Chersich S, Rejšek K, Vranová V, et al. Climate change impacts on the Alpine ecosystem: an overview with focus on the soil — a review. Journal of Forest Science, 2015, 61(11): 496–514
[52] Spohn M. Microbial respiration per unit microbial biomass depends on litter layer carbon-to-nitrogen ratio. Biogeosciences, 2015, 12(3): 817–823
[53] De Vries F T, Manning P, Tallowin J R B, et al. Abiotic drivers and plant traits explain landscape-scale patterns in soil microbial communities. Ecology Letters, 2012, 15(11): 1230–1239
[54] Tian Liming, Zhao Lin, Wu Xiaodong, et al. Soil moisture and texture primarily control the soil nutrient stoichiometry across the Tibetan grassland. Science of the Total Environment, 2018, 622/623: 192–202
[55] Liang Zhi, Olesen J E, Jensen J L, et al. Nutrient availability affects carbon turnover and microbial physiology differently in topsoil and subsoil under a temperate grassland. Geoderma, 2019, 336(7): 22–30
[56] Chen Leiyi, Liu Li, Mao Chao, et al. Nitrogen avai-lability regulates topsoil carbon dynamics after per-mafrost thaw by altering microbial metabolic effici-ency. Nature Communications, 2018, 9(1): 1–11
Spatial Pattern and Driving Factors of Soil Microbial Biomass Carbon in Grassland in Northern Agro-Pastoral Transition Zone
Abstract In order to explore spatial pattern of soil microbial biomass carbon (MBC) and its environmental driving forces in grassland surface layer (0‒10 cm) and subsurface layer (10‒20 cm) in the agro-pastoral transition zone in northern China, 456 soil samples (57 sample sites, two soil layers and four replicates) were selected for investigation from Inner Mongolia-Liaoning plain, Bashang on the north of Beijing, the northern foot of Yinshan Mountain and Ningxia-Shaanxi Loess Plateau. The results showed that the spatial pattern of MBC was consistent between the surface and subsurface layer, which increased with latitude and had no significant change with longitude and elevation. With the increase of degradation degree, the reduction of soil microbial biomass in the subsurface layer was smaller than that in the surface layer. The difference of soil pH between the two layers decreased with the degree of degradation. The difference in soil MBC between surface and subsurface soils was regulated by the change of soil pH. The smaller the difference in pH between the two soil layers, the smaller the difference in MBC. Climate, vegetation and soil factors could affect the spatial variation of MBC, and soil factors were the main driving forces for MBC at the regional scale. The main influencing factor of MBC was soil total carbon at surface layer, and soil total nitrogen at subsurface layer. In the context of climate change and the intensification of human activities, the results were useful for predicting the response of soil microorganisms at the regional scale and maintaining and restoring ecological functions of degraded grasslands.
Key words microbial biomass carbon (MBC); soil depth; agro-pastoral transition zone; grassland ecosystem; soil pH
doi: 10.13209/j.0479-8023.2021.003
国家自然科学基金(31972939, 31630009, 31670325)资助
收稿日期: 2020–02–26;
修回日期: 2020–04–03