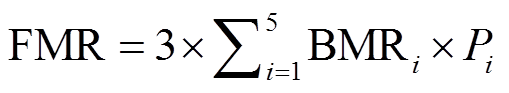 , (5)
, (5)摘要 针对湿地生态健康评价中的鸟类承载力评估问题,建立底栖动物去灰分干重(AFDW)的换算方法, 优化湿地鸟类体型分类标准。以深圳湾福田红树林湿地为例, 通过计算底栖动物总资源量、可支持水鸟总热值和水鸟种群的野外代谢率, 评估福田红树林湿地的水鸟承载力。结果表明: 1)福田红树林湿地底栖动物的总资源量具有明显的季节特征, 冬季(4.67×104kg)<春季(6.08×104kg)<夏季(8.00×104kg)<秋季(1.23×105kg), 以秋季为例, 不同生境的单位面积资源量为红树林植被区(89.22g/m2)>滩涂区(3.58g/m2)>基围鱼塘区(0.22g/m2); 2)湿地不同季节可支持水鸟的总热值为冬季(1.03×108kJ)<春季(1.36×108 kJ)<夏季(1.76×108 kJ)<秋季(2.70×108kJ); 3)福田红树林湿地水鸟种群野外代谢率为 467.27 kJ/d; 4)秋季、冬季和春季是候鸟迁徙期, 福田红树林湿地对水鸟的承载力分别为 6431, 2438 和 3235 只, 而实际观测数据高于研究结果, 说明当前福田红树林湿地对水鸟的承载力不足, 不能满足候鸟迁徙季节的鸟类食物需求, 湿地水鸟可能存在数量下降的风险。建议从加强红树林的保护与恢复重建、开展基围鱼塘生态修复和功能提升工程、强化鸟类和底栖动物的动态监测 3 个方面, 继续加大对福田红树林湿地的生态保护力度, 以期提高底栖动物总资源量, 进而提升红树林湿地的鸟类承载力。
关键词 深圳湾; 红树林湿地; 底栖动物资源量; 鸟类承载力
红树林湿地是热带、亚热带海岸带的重要湿地类型, 是地球上生产力最高的四大海洋生态系统之一, 其特殊的咸淡水交替环境使之成为动植物及微生物丰富的基因库[1]。红树林具有高光合率、高呼吸率和高归还率的特点, 为湿地底栖动物的生命活动提供能量来源, 丰富的底栖动物又为鸟类等高等动物提供充足的食物来源[2–3]。红树林湿地在粤港澳大湾区广泛分布, 是大湾区滨海湿地的重要组成部分[4]。生态健康评价是红树林监测管理的重要内容, 鸟类承载力评估是其中的重要环节。红树林湿地中的鸟类生境可分为 4 个部分: 红树林植被区、滩涂区、基围鱼塘区和农田稀树灌丛区[5]。滩涂区是水鸟觅食的主要区域, 涨潮时以游禽为主, 退潮时以涉禽(如鸻鹬类和鹭类等)为主[6]。红树林植被区是鹭类的栖息地和繁殖地以及一些秧鸡类的觅食地。基围鱼塘是水鸟另一个适宜的觅食地点, 调查显示, 20世纪 90 年代福田保护区基围鱼塘的破坏显著地影响鸻鹬类水鸟的数量[6]。农田稀树灌丛区是多种雀形目鸟类和其他陆鸟的栖息场所。
环境对生物的承载力表示环境所能承载的生物数量的范围, 主要受食物资源量、栖息地面积以及其他胁迫因子的影响。计算承载力时, 通常将食物供应量作为主要的考虑因素[7]。日消耗量模型(Daily Ration Model, DRM)是计算鸟类承载力的一种重要方法, 该模型定义生境中鸟类可获得的食物总生物量与每只鸟每日消耗的生物量的商即为该生境的鸟类承载力[8]。然而, 现有研究中对鸟类食量的直接观察较少, 多是根据林德曼效率原则进行换算[9–11]。Meire 等[12]通过分析涉禽去脂体重、基础代谢率以及日摄食量的关系, 评估河口区水鸟对底栖动物的消费量和承载力。底栖动物是红树林湿地水鸟的主要食物来源, 其资源量与水鸟丰度和多样性等指标之间有很好的相关性[13]。现有研究主要根据湿地底栖动物提供的总能量、水鸟每天野外代谢消耗的能量以及迁徙时的滞留时间, 计算湿地所能支持水鸟最大种群数量[8,14]。
Custard 等[8]在评价鸟类承载力时, 主要使用底栖动物的去灰分干重数据, 由于数据获取难度较大, 影响评价的可行性; 在考虑鸟类的体型种群结构时, 仅将体型划分为大、中、小 3 类, 影响鸟类种群综合代谢率计算的准确性。
本研究构建底栖动物“湿重–干重–去灰分干重”的换算方法, 能够降低数据获取难度, 提高评估效率。同时, 细化基于体型的鸟类种群结构划分标准, 有效地提高鸟类承载力评价方法的准确性。本文基于深圳湾福田红树林湿地面积和底栖动物生物量湿重数据以及底栖动物“湿重–干重–去灰分干重”的换算方法, 计算湿地底栖动物总资源量和可支持水鸟的总热值情况。基于优化的鸟类种群结构划分标准, 对湿地水鸟进行种群结构的再构建以及种群野外代谢率的计算。本研究进一步评价福田红树林湿地对水鸟的承载力, 并提出相应的鸟类承载力提升对策。
深圳福田红树林国家级自然保护区是全国唯一地处城市腹地的国家级自然保护区, 位于深圳湾东北部(113°45′E, 22°32′N), 与香港米埔红树林湿地隔海相望, 属南亚热带季风气候, 年平均气温为 22℃, 年均降水量为 1927mm, 年均相对湿度为 79%[15]。深圳湾福田红树林湿地内水鸟的主要觅食场所包括滩涂区、红树林植被区和基围鱼塘区。该区域总面积为 337.95hm2, 其中滩涂区面积为 137.79hm2, 红树林植被区面积为 132.84hm2, 基围鱼塘区面积为67.32hm2[16]。
本研究使用的底栖动物生物量(fresh weight, FW)数据和鸟类统计数据来自《广东内伶仃–福田国家级自然保护区福田红树林生物多样性与生态环境监测报告(2017 年度)》, 简称《监测报告(2017年度)》。底栖动物的生物量依据《海洋调查规范》、《海洋底栖生物研究方法》和《红树林生态检测技术规程》, 于 2017 年 2—11 月进行春、夏、秋、冬(分别为 5 月、8 月、11 月和 2 月)周年采样, 采样点包括观鸟亭、凤塘河、沙咀和基围鱼塘。采用样点法, 通过野外观测获取鸟类统计数据, 包括53 种水鸟(分属 6 目 10 科)的各月份数量及各种类数量, 监测点包括凤塘河口、沙嘴鱼塘和新洲河口。
以日消耗量模型和林德曼效率原则为理论基础[11], 计算福田红树林湿地不同季节的水鸟承载力。主要包括底栖动物总资源量、湿地可支持水鸟总热值、鸟类种群野外代谢率和湿地水鸟承载力。
1.3.1底栖动物总资源量
通过文献检索, 收集多毛纲、双壳纲、腹足纲、甲壳纲和硬骨鱼纲底栖动物的含水率和去灰分干重百分比数据, 基于湿重数据计算深圳湾福田红树林湿地底栖动物的去灰分干重生物量。结合福田红树林植被区、滩涂区和基围鱼塘区的面积, 计算底栖动物的总资源量。主要步骤如下。
1)计算底栖动物去灰分干重生物量。基于不同生境类型中底栖动物的湿重数据, 计算福田红树林湿地底栖动物的去灰分干重生物量(ash free dry weight, AFDW)(g/m2):
AFDW = FW × (1 – w0) × w1, (1)
式中, FW 为底栖动物的湿重生物量(g/m2), w0 为底栖动物的含水率(%), w1 为底栖动物干重中的去灰分干重百分比(%)。
鉴于福田红树林湿地中鸟类觅食区主要在红树林植被区、滩涂区和基围鱼塘区, 依此对原始采样点进行基于觅食场所的分类和再计算, 将底栖动物生物量的平均值作为各觅食场所的底栖动物生物量。底栖动物基于纲分类的含水率(w0)通过文献检索获取: 双壳纲为 80.0%[17–18], 腹足纲为 80.7%[18], 硬骨鱼纲为 81.5%[19], 多毛纲为 80.2%[20], 甲壳纲取虾和蟹的平均值为 78.8% (虾为 76.3%[21]、蟹为81.2%[22])。底栖动物基于纲分类的去灰分干重百分比(w1)通过文献获取: 多毛纲为 81.5% (用 0.5mm 和0.2mm 网孔采集条件下的数据平均值代替)[23], 双壳纲为 75.0%[23], 腹足纲为 75.4%[24], 甲壳纲为75.0%[25], 硬骨鱼纲为84.6%[26]。
2)计算底栖动物总资源量。红树林湿地的底栖动物总资源量为湿地中所有底栖动物的去灰分干重的总和, 计算公式如下:
Ci=Si×AFDWi×10, (2)
C =C1+C2+C3, (3)
式中, C1, C2 和 C3 分别为红树林植被区、滩涂区和基围鱼塘区的底栖动物资源量(kg), Si 为各生境类型的面积(hm2), AFDWi 为各生境类型内的单位面积底栖动物去灰分干重(g/m2), C 为红树林湿地中的底栖动物总资源量(kg)。
1.3.2湿地可支持水鸟总热值
基于红树林湿地中鸟类的实际种群结构, 将所调查的全部鸟类视为红树林湿地碎屑食物链中的一个营养级, 底栖动物为湿地鸟类直接且唯一的下一营养级, 反之亦然。根据林德曼效率原则, 湿地可支持水鸟的总热值约为底栖动物总热值的 1/10[10]。计算公式如下:
Q = C×1000×F×10%, (4)
式中, Q为湿地可支持水鸟总热值(kJ), 潮间带底栖动物的热值F=22 kJ/g[27]。
1.3.3鸟类种群野外代谢率
以 2017 年的福田红树林湿地鸟类统计数据为基础, 根据水鸟体长(数据来自文献[28], 部分取中值), 将监测的水鸟划分为大型(体长>50cm)、较大型(40cm<体长≤50cm)、中型(30cm<体长≤40cm)、较小型(20cm<体长≤30cm)和小型(体长<20cm)5 个等级, 并基于体长, 对鸟类的种群结构进行再构建。
根据不同体长水鸟的代谢率, 计算红树林湿地水鸟种群的野外代谢率(field metabolic rate, FMR) (kJ/d), 公式如下:
, (5)式中, BMRi 为各体型鸟类的平均基础代谢率(kJ/ d)[12]: 308.50(大型)、195.00(较大型)、125.00(中型)、113.00(较小型)和53.00(小型); Pi为各体型鸟类在湿地鸟类种群中的占比(%); 常数 3 的含义为FMR 按各鸟类种群中个体平均综合基础代谢率的 3倍计算[9]。
1.3.4湿地水鸟承载力
各季节水鸟承载力(只)及承载力密度计算公式如下:
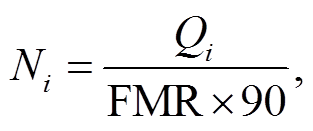 (6)
(6) (7)
(7)
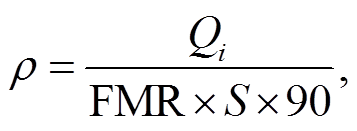 (8)
(8)
式中, Ni 为各季节的水鸟承载力(只), Ni′为以鸟日计的各季节单位面积的水鸟承载力(鸟日/hm2), ρ 为各季节单位面积的水鸟承载力(只/hm2), Qi 为各季节的可支持水鸟总热值(kJ), S 为红树林湿地生境总面积(hm2), 常数 90 的含义为各季节水鸟的停留时间按 90 天计算。
根据最优觅食理论(Optimal Foraging Theory)[29], 动物在觅食时, 期望用最小的成本获得最大的收益。野生动物更倾向于在生物量更高的环境中觅食, 因此底栖动物的生物量和分布可能会影响鸟类对觅食场所的选择和食物构成。如图 1 所示, 深圳湾福田红树林湿地 3 种鸟类觅食区(红树林植被区、滩涂区和基围鱼塘区)底栖动物生物量(FW)的季节变化特征为冬季<春季<夏季<秋季, 最高的底栖动物生物量出现在秋季的红树林植被区(FW= 586.34g/m2), 这一季节性分布特征与周细平等[30]在九龙江口海门岛红树林湿地的调查结果类似, 该研究发现大型底栖动物生物量最高的季节为秋季(FW =419.48g/m2)。
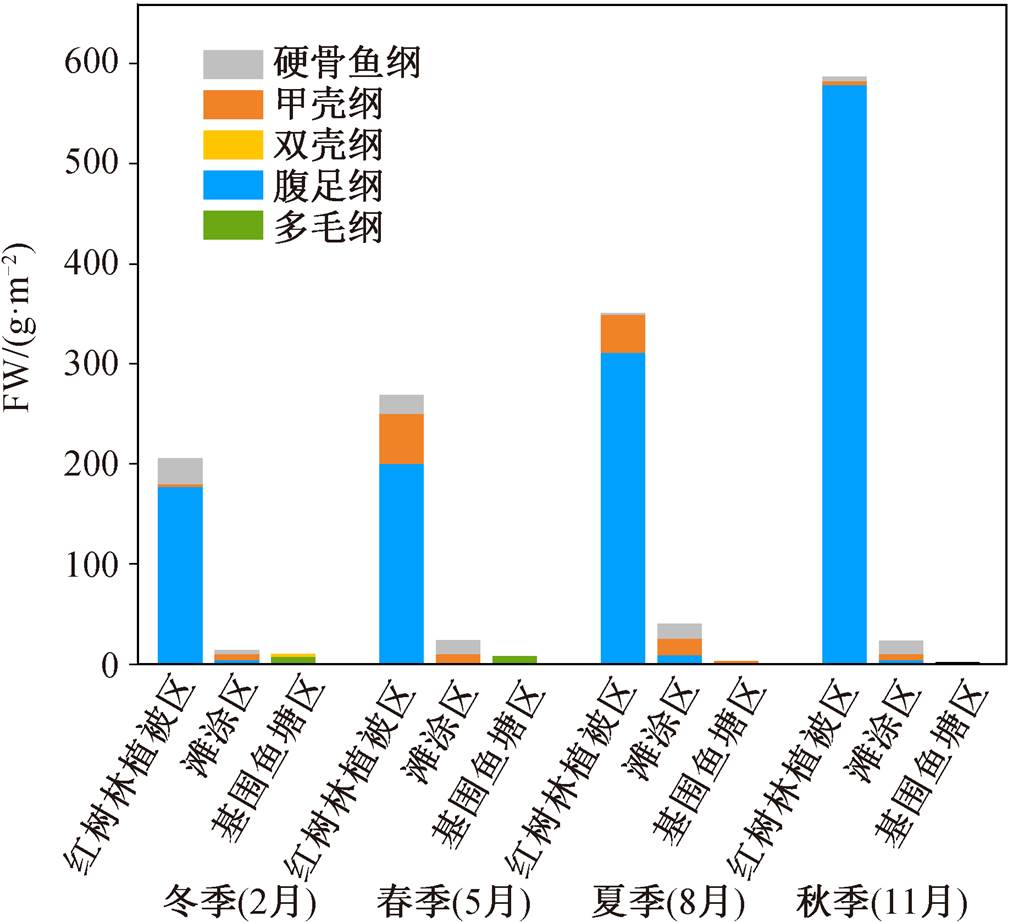
图1 福田红树林湿地不同季节和生境底栖动物生物量特征
Fig. 1 Biomass characteristics of benthos in different seasons and habitats of Futian mangrove wetland
这种季节变化规律可能与底栖动物的生长规律以及当地的鸟类活动规律有关。由于鸟类与底栖动物之间存在捕食与被捕食的关系, 福田红树林湿地鸟类的活动时间集中在 11 月至次年 4 月, 在这期间对底栖动物的消耗使其生物量降低, 由于鸟类的迁徙行为, 底栖动物资源量又逐步恢复, 直至秋季达到最大量。对香港米埔红树林湿地的相关研究也支持这一观点, 该研究在对 3 种主要水鸟(涉禽类、鸭类和鸥类)周年食物消耗量的观察中发现明显的季节性特点, 食物消耗集中在 11 月至次年 4 月(冬季), 且在 1 月达到最高峰[6]。九段沙湿地位于长江口区域, 春秋迁徙季节是主要的水鸟活动季节[31], 其底栖动物的季节分布与纬度较低的福田红树林湿地和九龙江海门岛红树林湿地呈现不同的规律, 春季(AFDW = 3.13~100.17g/m2)大于秋季(AFDW = 17.73 ~47.59g/m2)[30]。
以秋季为例, 总底栖动物生物量(以湿重计)在不同生境类型间的分布特征为红树林植被区(586.34g/m2) > 滩涂区(23.11g/m2) > 基围鱼塘区(1.39g/m2), 但不同种类底栖动物之间存在差异。全年范围内, 腹足纲和多毛纲分别为红树林植被区和基围鱼塘区主要的底栖动物种类, 但不同种类底栖动物具有不同的分布规律, 可能与红树林植被区独特的环境特征有关。红树植物的隐蔽作用使得红树植被区的物理环境(温度、光照、空气湿度及海浪的冲击程度等)更加稳定温和, 底栖动物受到的环境胁迫较小(比如腹足类通常聚集在荫蔽和潮湿的红树植物周围的地表或地面根中[32]), 红树林植被区软体动物物种数量和生物量都高于滩涂区[33]; 红树植物残片在底质中的分解和淋溶, 提高了红树林植被区底质有机质的含量, 并为底栖动物提供营养来源[34]。这些因素决定了红树林植被区更适宜底栖动物的生存, 因而拥有较高的底栖动物生物量。
福田红树林湿地的底栖动物单位面积去灰分干重生物量和总资源量如表 1 所示。福田红树林湿地的总资源量在秋季最大(1.2×105kg), 在春季最小(4.7×104kg)。经换算后, 秋季底栖动物的去灰分干重生物量在不同生境类型间的分布趋势总体上与湿重生物量类似: 红树林植被区(85.42g/m2) > 滩涂区(3.58g/m2) > 基围鱼塘区(0.22g/m2)。总之, 红树林植被区的资源量最高, 鸟类的主要食物类型可能是腹足纲的底栖动物。福田红树林湿地的底栖动物生物量与九段沙湿地(春季 23.13~100.17g/m2, 秋季17.73~47.59g/m2, AFDW)[11]相近。红树林植被区的生物量与九龙江红树林湿地的底栖动物年平均生物量(77.58g/m2, AFDW)[30]相近, 滩涂区和基围鱼塘区的生物量远低于此值。鸟类生境的退化使得涠洲岛湿地潮间带的生物量(春季 4.2±3.9g/m2, 秋季 5.4± 5.0g/m2, AFDW)远低于福田红树林湿地。中街山列岛岩礁潮间带底栖动物平均湿重生物量达到9651.97±3911.92g/m2 [35], 江苏北部的东西连岛、车牛山岛、达山岛及平山岛为 4150.2g/m2[36], 这些潮间带环境中的底栖动物生物量远高于福田红树林湿地。因此, 福田红树林湿地的底栖动物生物量仍有很大的提升空间。
表1 福田红树林各生境类型面积和不同季节底栖动物资源量(以去灰分干重计)
Table 1 Area and food mass of each habitat in Futian mangrove wetland during different seasons (AFDW)

季节红树林植被区滩涂区基围鱼塘区总计AFDW/(g·m–2)C/kgAFDW/(g·m–2)C/kgAFDW/(g·m–2)C/kgAFDW/(g·m–2)C/kg 冬季30.12415012.1329391.57216733.8246607 春季39.96550573.6950911.23169744.8861845 夏季51.50709686.1985310.3953158.0880029 秋季85.421176943.5849360.2230889.22122938 面积[16]137.79 hm2132.84 hm267.32 hm2337.95 hm2
福田红树林湿地各生境不同季节的可支持水鸟总热值情况如图 2 所示。可支持水鸟总热值的季节特征与总资源量的趋势一致: 冬季(1.03×108kJ) <春季(1.36×108kJ) <夏季(1.76×108kJ) <秋季(2.70×108kJ)。生境分布特征也与总资源量分布特征一致, 以总热值最高的秋季为例, 基围鱼塘区(6.78×105kJ) < 滩涂区(1.09×107kJ) <红树林植被区(2.59×108kJ)。为了保证评价结果的可获得性, 本研究对所有底栖动物采用相同的热值(F=22kJ/g[27])。作为评估湿地总热值的关键性数据, 底栖动物热值对评价结果的准确性有较大影响。通过实验确定底栖动物的实际热值情况, 能够增强评价结果的合理性和可靠性, 但也会增加工作量, 从而影响评价效率, 在评价过程中应根据实际需求做出取舍。
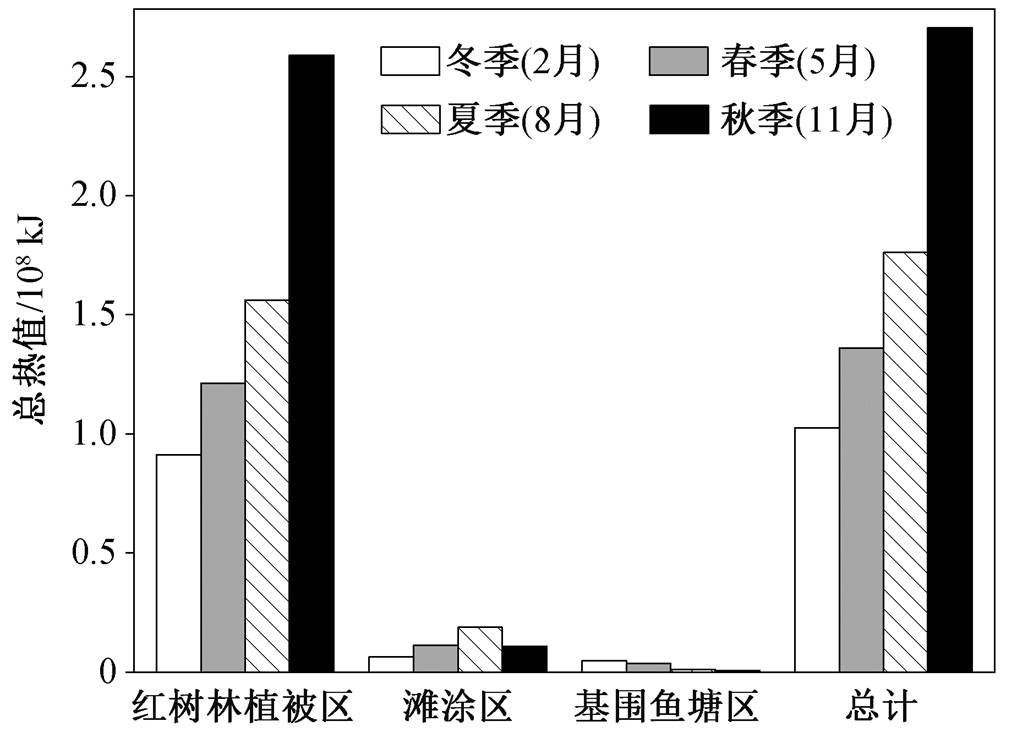
图2 福田红树林各生境类型不同季节可支持水鸟总热值
Fig. 2 Maximum calories for shorebirds in each habitat of Futian mangrove wetland during different seasons
根据福田红树林湿地鸟类迁徙规律[37], 11 月至次年 4 月鸟类种类和数量较多(即冬候鸟活动期)。因此, 对该区鸟类承载力的分析以秋季、冬季和春季为主。同时, 该区鸟类种群结构以水鸟为主(约99%, 数据来自《监测报告(2017 年度)》), 因此以水鸟作为主要研究对象。福田红树林湿地各种属水鸟的居留型特征、体型分类以及种群占比见附录。基于体型分类的种群结构再构建及综合基础代谢率折算结果如表 2 所示。水鸟体型以较大型(40cm <体长≤ 50cm)和较小型(20cm < 体长 ≤ 30cm)为主, 占比分别为 35.66%和 29.46%, 水鸟种群的综合基础代谢率为 155.76 kJ/d, 野外代谢率为 467.27 kJ/d。
进一步地, 根据福田红树林湿地可支持水鸟总热值、鸟类的野外综合基础代谢率以及迁徙期滞留天数, 得到福田红树林湿地的水鸟承载力及承载力密度(表 3)。深圳湾福田红树林湿地对水鸟的承载力(秋季、冬季和春季)分别为 6431, 2438 和 3235只, 承载力密度分别为 19.0, 7.2 和 9.6 只/hm2, 相当于 1712.7, 649.3 和 861.6 鸟日/hm2。根据《监测报告(2017 年度)》中基于样点法的野外观测获得的福田红树林湿地水鸟数量周年变化情况, 水鸟数量在 448 只(8 月份)~8994 只(12 月份)之间。一年中湿地对水鸟的最大承载力(6431 只, 秋季)仍小于湿地在 12 月份水鸟的个体数量(8994 只), 表明当前福田红树林湿地对水鸟的承载力不足, 尚不能满足候鸟迁徙季节的鸟类食物需求, 湿地水鸟存在数量下降的风险。涠洲岛湿地对鸻鹬类水鸟的承载力, 春秋季分别为 874~1236 只和 1001~1302 只; 承载力密度(只/hm2)为春季库塘(10.7)>沼泽(5.6)>潮间带(4.8), 秋季沼泽(6.8) >库塘(4.3) >潮间带(3.6); 承载力相当于 568~739 鸟日/hm2 [9]。长江口九段沙的春季以及秋季鸟类承载力分别为 174.7 和 138.8 鸟日/hm2[11]。美国南旧金山湾盐田湿地的承载力为 9443±1649 鸟日/hm2 [12]。可见, 与处于生境退化时期的涠洲岛湿地相比, 福田红树林湿地对水鸟的承载力处于较高水平, 但与一些其他的湿地鸟类生境相比, 仍有较大的提升空间。
表 2 福田红树林湿地水鸟种群结构和综合基础代谢率折算
Table 2 Community component and synthetic BMR of the shorebirds in Futian mangrove wetland
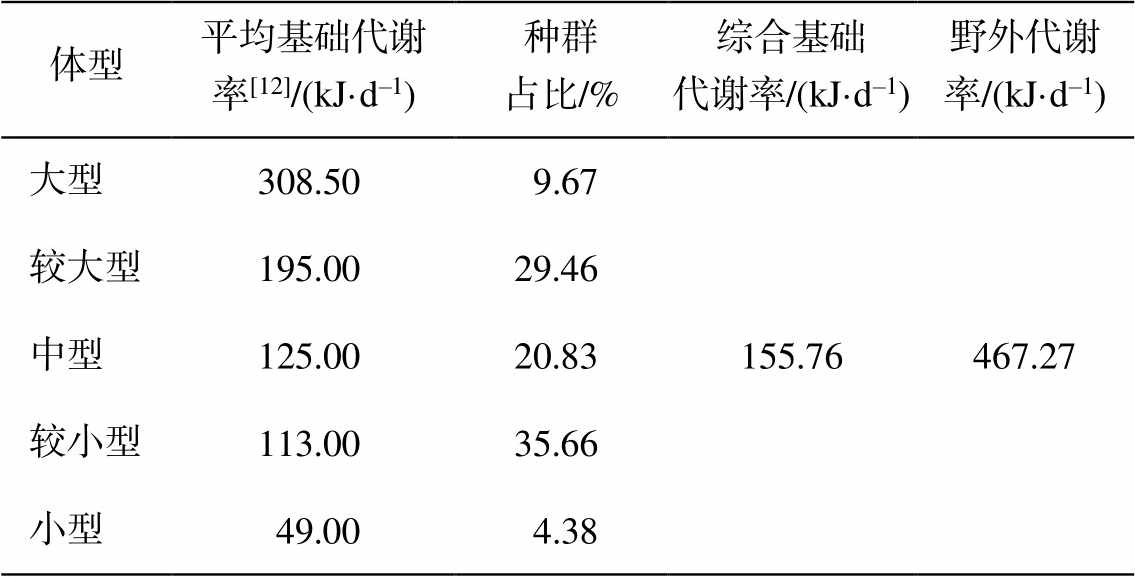
体型平均基础代谢率[12]/(kJ·d–1)种群占比/%综合基础代谢率/(kJ·d–1)野外代谢率/(kJ·d–1) 大型308.509.67155.76467.27 较大型195.0029.46 中型125.0020.83 较小型113.0035.66 小型49.004.38
表3 福田红树林湿地越冬期水鸟的承载力
Table 3 Carrying capacity of overwintering shorebirds in Futian mangrove wetland
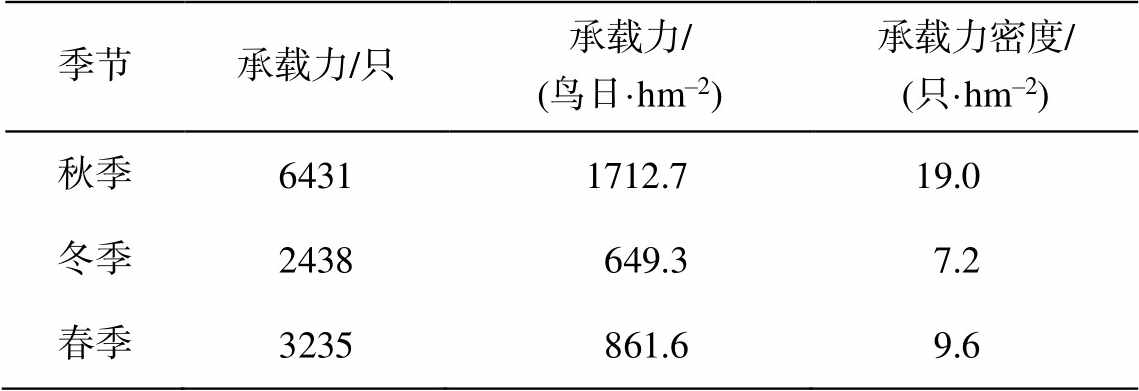
季节承载力/只承载力/ (鸟日·hm–2)承载力密度/ (只·hm–2) 秋季64311712.7 19.0 冬季2438649.3 7.2 春季3235861.6 9.6
1)加强红树林植被的保护与恢复重建。福田红树林湿地的红树林植被区拥有最高的底栖生物资源量, 在承载湿地鸟类方面起着重要的作用。一方面, 应积极维持和加强红树林植被的保护, 遏制红树林生态功能退化; 另一方面, 对已破坏的红树林进行植被恢复和重建, 适度增加红树植物面积。
2)开展基围鱼塘生态修复和功能提升工程。基围鱼塘区也是鸟类觅食和栖息的重要场所[38–39], 本研究的评估结果表明, 福田红树林湿地中的基围鱼塘区底栖动物资源量较低, 对鸟类承载力的贡献较低。已开展的基围鱼塘生态功能提升工程已取得明显的效果, 如 2000 年对福田沙嘴鱼塘的改造提升了鸟类的密度、丰度和种群多样性指数, 表明鱼塘的定向改造对其生态功能和生态承载力的恢复和提高有积极的作用[40]。因此, 针对福田红树林湿地基围鱼塘生态功能低下的问题, 可进一步实施生态修复和功能提升工程, 如通过控制水位的方式, 借助潮汐, 为水鸟带来丰富的饵料, 提高底栖动物和鱼类等的资源量; 通过堤埂修整、合理配置植物的方式提升鱼塘的生境多样性; 通过引入鸟类实时在线监测等智能监测技术, 及时有效地开展修复后评价, 检验和评价工程效果。
3)加强鸟类和底栖动物的动态监测。目前, 福田红树林湿地对鸟类的承载力不足, 存在一定的生态健康风险。对生态系统主要组分的系统调查和动态监测是评价湿地生态健康状况的基础[41]。因此, 应加强对鸟类和底栖动物的资源调查和动态监测, 以便实时掌握底栖动物和鸟类的健康状况, 及时识别风险, 并做出相应的应对措施。
本研究针对湿地生态健康评价中的鸟类承载力评估问题, 建立底栖动物去灰分干重(AFDW)的换算方法, 优化湿地鸟类体型分类标准。本文以深圳湾福田红树林湿地为例, 通过计算底栖动物总资源量、可支持水鸟总热值以及水鸟种群的野外代谢率, 来评估福田红树林湿地的水鸟承载力, 得到如下结论。
1)福田红树林湿地底栖动物的总资源量具有季节性特征: 冬季(4.67×104kg)<春季(6.08×104kg)<夏季(8.00×104kg)<秋季(1.23×105kg)。以资源量最高的秋季为例, 不同生境的单位面积资源量存在差异: 红树林植被区(89.22g/m2)>滩涂区(3.58g/m2)>基围鱼塘区(0.22g/m2)。底栖动物以腹足纲为主。
2)福田红树林湿地不同季节可支持水鸟的总热值为冬季(1.03×108kJ)<春季(1.36×108kJ)<夏季(1.76×108kJ)<秋季(2.70×108kJ)。
3)深圳湾福田红树林湿地水鸟种群的综合基础代谢为 155.76kJ/d, 野外基础代谢率为 467.27kJ/d。体型类别以较大型(40cm<体长≤50cm)和较小型(20cm<体长≤ 30cm)为主, 占比分别为 35.66%和29.46%。
4)冬季、春季和秋季是候鸟的迁徙期, 深圳湾福田红树林湿地对水鸟的承载力分别为 2438, 3235和 6431 只, 承载力密度分别为 7.2, 9.6 和 19.0 只/hm2, 相当于 649.3, 861.6 和 1712.7 鸟日/hm2。2017年基于样点法的野外观测结果表明, 福田红树林湿地水鸟数量最多时可达 8994 只, 当前湿地的水鸟承载力不足。
建议从加强红树林的保护与恢复重建、开展基围鱼塘的生态修复和功能提升工程以及强化鸟类和底栖动物的动态监测 3 个方面, 继续加大对福田红树林湿地的生态保护力度, 以期提高底栖动物总资源量, 进而提升湿地的鸟类承载力。
参考文献
[1] 宋晓军, 林鹏. 福建红树林湿地鸟类区系研究. 生态学杂志, 2002, 21(6): 5–10
[2] 范航清, 韦受庆, 陈坚. 广西红树林区经济动物的行为生态及其生态养殖的初步设计. 广西科学院学报, 1993, 9(2): 104–110
[3] 林鹏, 陈荣华. 红树林有机碎屑在河口生态系统中的作用. 生态学杂志, 1991(2): 45–48, 64
[4] 于凌云, 林绅辉, 焦学尧, 等. 粤港澳大湾区红树林湿地面临的生态问题与保护对策. 北京大学学报(自然科学版), 2019, 55(4): 782–790
[5] 王勇军, 刘治平, 陈相如. 深圳福田红树林冬季鸟类调查. 生态科学, 1993, 12(2): 77–84
[6] 林鹏. 中国红树林生态系. 北京: 科学出版社, 1997
[7] Goss-custard J D, Stillman R A, West A D, et al. Carrying capacity in overwintering migratory birds. Biological Conservation, 2002, 105(1): 27–41
[8] Custard J D G, Stillman R A, Caldow R W G, et al. Carrying capacity in overwintering birds: when are spatial models needed?. Journal of Applied Ecology, 2003, 40(1): 176–187
[9] 莫竹承, 孙仁杰, 陈骁, 等. 涠洲岛湿地对鸻鹬类水鸟的承载力评估. 广西科学, 2018, 25(2): 181–188
[10] 陈进树. 浅析林德曼效率的适用前提和对象. 中国科技信息, 2010(17): 24
[11] 葛振鸣, 周晓, 施文彧, 等. 九段沙湿地鸻形目鸟类迁徙季节环境容纳量. 生态学报, 2007, 27(1): 90–96
[12] Meire P M, Schekkerman H, Meininger P L. Consum-ption of benthic invertebrates by waterbirds in the Oosterschelde estuary, SW Netherlands. Hydrobiolo-gia, 1994, 282(1): 525–546
[13] 林清贤. 闽南沿海红树林区鸟类及其与大型底栖动物相关关系研究[D]. 厦门: 厦门大学, 2003
[14] Brand L A, Takekawa J Y, Shinn J, et al. Effects of wetland management on carrying capacity of diving ducks and shorebirds in a coastal estuary. Waterbirds, 2014, 37(1): 52–67
[15] 李瑞利, 柴民伟, 邱国玉, 等. 近 50 年来深圳湾红树林湿地 Hg、Cu 累积及其生态危害评价. 环境科学, 2012, 33(12): 4276–4283
[16] 陈志云, 牛安逸, 徐颂军, 等. 基于地学信息图谱的深圳湾湿地景观变化分析. 林业科学, 2018, 54(3): 168–176
[17] Usero J, Morillo J, Gracia I. Heavy metal concentra-tions in molluscs from the Atlantic coast of southern Spain. Chemosphere, 2005, 59(8): 1175–1181
[18] Wei Y H, Zhang J Y, Zhang D W, et al. Metal concentrations in various fish organs of different fish species from Poyang Lake, China. Ecotoxicology and Environmental Safety, 2014, 104: 182–188
[19] Yu T, Zhang Y, Hu X N, et al. Distribution and bioaccumulation of heavy metals in aquatic organisms of different trophic levels and potential health risk assessment from Taihu lake, China. Ecotoxicology and Environmental Safety, 2012, 81: 55–64
[20] 刘天红, 于道德, 李红艳, 等. 东营养殖双齿围沙蚕营养成分分析及膳食营养评价. 水产科学, 2017, 36(2): 160–166
[21] 张军文, 郑晓伟, 沈建, 等. 南极磷虾干燥特性的研究. 极地研究, 2018, 30(2): 186–191
[22] 鲁丹, 俞琰垒, 张虹. 梭子蟹中硒形态分析及其分布研究. 食品研究与开发, 2016, 37(13): 109–112
[23] Widbom B. Determination of average individual dry weights and ash-free dry weights in different sieve fractions of marine meiofauna. Marine Biology, 1984, 84(1): 101–108
[24] Bernardini V, Solimini A G, Carchini G. Application of an image analysis system to the determination of biomass (ash free dry weight) of pond macroinver-tebrates. Hydrobiologia, 2000, 439(1/3): 179–182
[25] Horn S, de la Vega C. Relationships between fresh weight, dry weight, ash free dry weight, carbon and nitrogen content for selected vertebrates. Journal of Experimental Marine Biology and Ecology, 2016, 481: 41–48
[26] Lappalainen A, Kangas P. Littoral benthos of the Northern Baltic Sea II. interrelationships of wet, dry and ash-free dry weights of macrofauna in the Tvä-rminne Area. Internationale Revue der gesamten Hy-drobiologie und Hydrographie, 1975, 60(3): 297–312
[27] Zwarts L, Blomert A M. Selectivity of whimbrels fee-ding on fiddler-crabs explained by component specific digestibilities. Ardea, 1990,78(1/2): 193–208
[28] 尹琏, 费嘉伦, 林超英. 中国香港及华南鸟类野外手册. 长沙: 湖南教育出版社, 2017
[29] Macarthur R H, Pianka E R. On optimal use of a patchy environment. The American Naturalist, 1966, 100: 603–609
[30] 周细平, 徐帅良, 吴培芳, 等. 九龙江口海门岛红树林湿地大型底栖动物群落生态研究. 应用海洋学学报, 2019, 38(1): 21–29
[31] 葛振鸣. 长江口滨海湿地迁徙水禽群落特征及生境修复策略[D]. 上海: 华东师范大学, 2007
[32] 刘荣成. 中国惠安洛阳江红树林. 北京: 中国林业出版社, 2010
[33] 黄雅琴, 李荣冠, 江锦祥. 泉州湾洛阳江红树林自然保护区潮间带软体动物多样性及分布. 海洋科学, 2011, 35(10): 110–116
[34] Fratini S, Cannicci S, Vannini M. Feeding clusters and olfaction in the mangrove snail Terebralia palus-tris (Linnaeus) (Potamididae: Gastropoda). Journal of Experimental Marine Biology and Ecology, 2001, 261 (2): 173–183
[35] 姚海峰, 薛巍, 邹广明, 等. 中街山列岛岩礁潮间带春季大型底栖动物的群落结构. 安徽农业科学, 2012, 40(17): 9282–9284
[36] 张虎, 郭仲仁, 刘培廷. 江苏省海岛潮间带底栖生物分布特征. 南方水产, 2010, 6(4): 49–55
[37] 陈志鹏, 胡柳柳, 王皓, 等. 深圳福田红树林保护区水鸟调查及种群变化研究. 资源节约与环保, 2016(12): 163–167
[38] 王勇军, 昝启杰, 常弘. 深圳福田红树林湿地鹭科鸟类群落生态研究. 中山大学学报(自然科学版), 1999, 38(2): 86–90
[39] 陈桂珠, 王勇军, 黄乔兰. 深圳福田红树林鸟类自然保护区生物多样性及其保护研究. 生物多样性, 1997, 5(2): 104–111
[40] 王勇军, 徐华林, 昝启杰. 深圳福田鱼塘改造区鸟类监测及评价//第二届全国复合生态与循环经济学术讨论会. 合肥, 2005: 352–356
[41] 陈彬. 基于多源数据的鄱阳湖候鸟栖息地动态监测方法研究[D]. 北京: 中国科学院研究生院, 2015
附录 2017年福田红树林湿地水鸟体长和种群占比
Appendix Body length and community ratio of shorebirds in Futian mangrove wetland in 2017
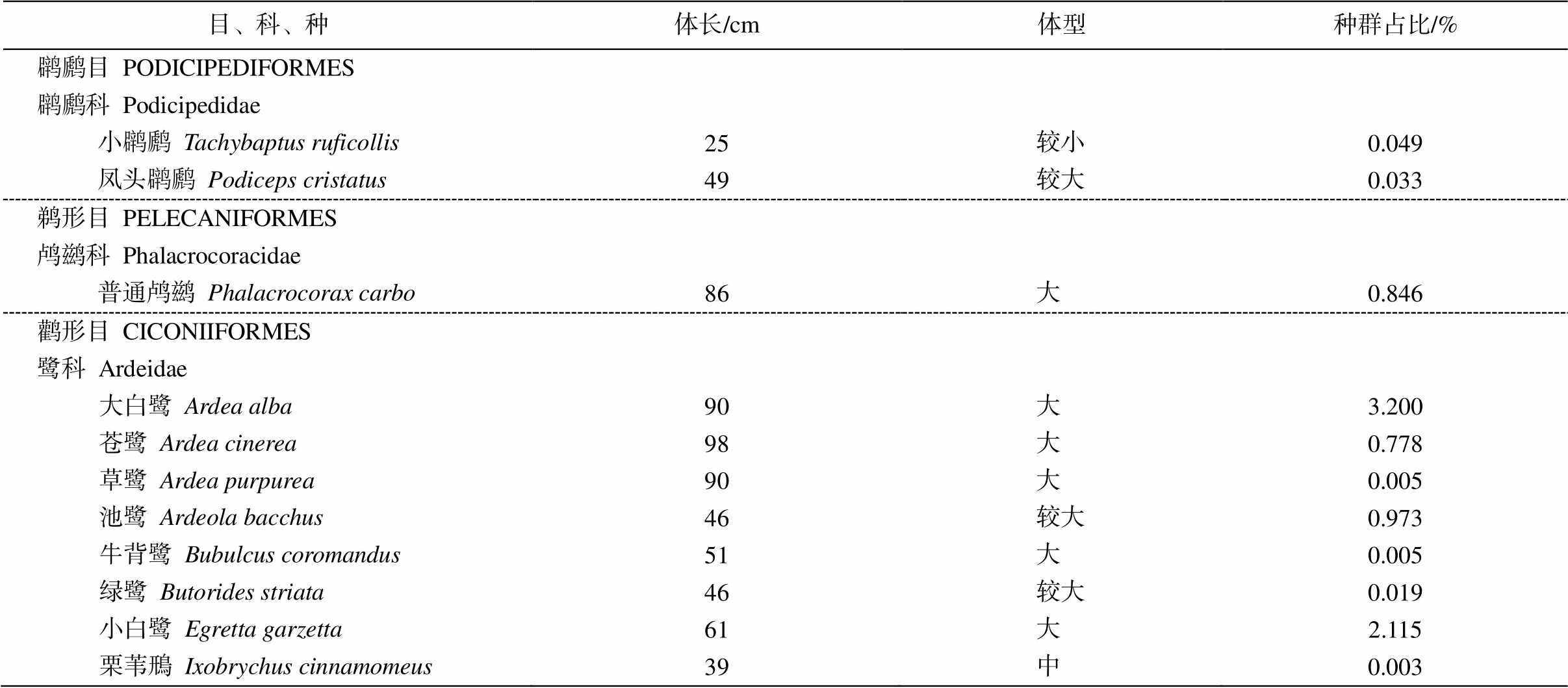
目、科、种体长/cm体型种群占比/% 䴙䴘目 PODICIPEDIFORMES 䴙䴘科 Podicipedidae 小䴙䴘Tachybaptus ruficollis25较小0.049 凤头䴙䴘Podiceps cristatus49较大0.033 鹈形目 PELECANIFORMES 鸬鹚科 Phalacrocoracidae 普通鸬鹚Phalacrocorax carbo86大0.846 鹳形目 CICONIIFORMES 鹭科 Ardeidae 大白鹭Ardea alba90大3.200 苍鹭Ardea cinerea98大0.778 草鹭Ardea purpurea90大0.005 池鹭Ardeola bacchus46较大0.973 牛背鹭Bubulcus coromandus51大0.005 绿鹭Butorides striata46较大0.019 小白鹭Egretta garzetta61大2.115 栗苇鳽Ixobrychus cinnamomeus39中0.003
续表
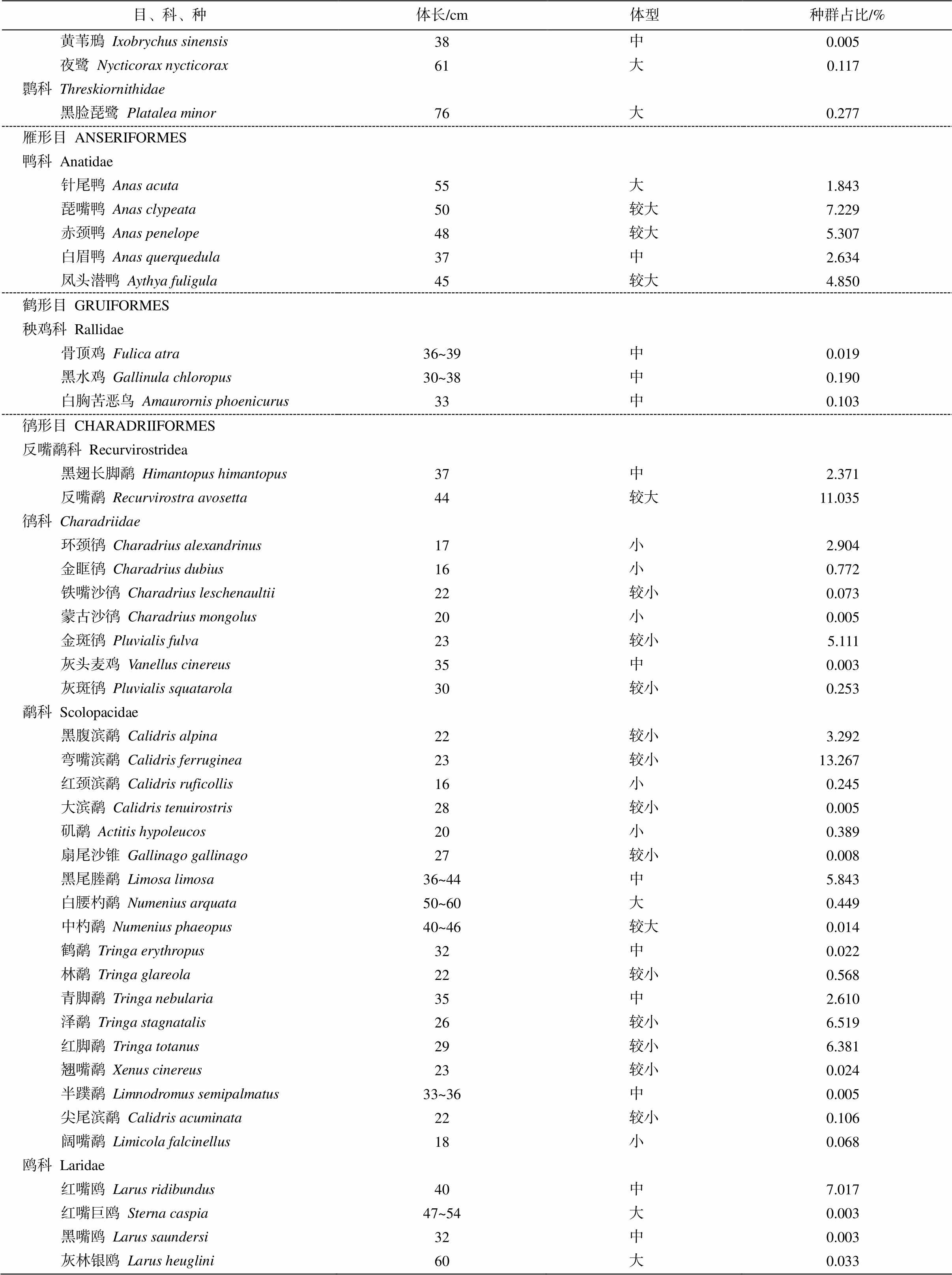
目、科、种体长/cm体型种群占比/% 黄苇鳽Ixobrychus sinensis38中0.005 夜鹭Nycticorax nycticorax61大0.117 鹮科Threskiornithidae 黑脸琵鹭 Platalea minor76大0.277 雁形目 ANSERIFORMES 鸭科 Anatidae 针尾鸭Anas acuta55大1.843 琵嘴鸭Anas clypeata50较大7.229 赤颈鸭Anas penelope48较大5.307 白眉鸭Anas querquedula37中2.634 凤头潜鸭Aythya fuligula45较大4.850 鹤形目 GRUIFORMES 秧鸡科 Rallidae 骨顶鸡Fulica atra36~39中0.019 黑水鸡 Gallinula chloropus30~38中0.190 白胸苦恶鸟 Amaurornis phoenicurus33中0.103 鸻形目 CHARADRIIFORMES 反嘴鹬科 Recurvirostridea 黑翅长脚鹬 Himantopus himantopus37中2.371 反嘴鹬 Recurvirostra avosetta44较大11.035 鸻科Charadriidae 环颈鸻Charadrius alexandrinus17小2.904 金眶鸻Charadrius dubius16小0.772 铁嘴沙鸻Charadrius leschenaultii22较小0.073 蒙古沙鸻Charadrius mongolus20小0.005 金斑鸻Pluvialis fulva23较小5.111 灰头麦鸡Vanellus cinereus35中0.003 灰斑鸻Pluvialis squatarola30较小0.253 鹬科 Scolopacidae 黑腹滨鹬Calidris alpina22较小3.292 弯嘴滨鹬 Calidris ferruginea23较小13.267 红颈滨鹬 Calidris ruficollis16小0.245 大滨鹬 Calidris tenuirostris28较小0.005 矶鹬 Actitis hypoleucos20小0.389 扇尾沙锥 Gallinago gallinago27较小0.008 黑尾塍鹬Limosa limosa36~44中5.843 白腰杓鹬 Numenius arquata50~60大0.449 中杓鹬 Numenius phaeopus40~46较大0.014 鹤鹬Tringa erythropus32中0.022 林鹬Tringa glareola22较小0.568 青脚鹬Tringa nebularia35中2.610 泽鹬Tringa stagnatalis26较小6.519 红脚鹬 Tringa totanus29较小6.381 翘嘴鹬Xenus cinereus23较小0.024 半蹼鹬 Limnodromus semipalmatus33~36中0.005 尖尾滨鹬Calidris acuminata22较小0.106 阔嘴鹬Limicola falcinellus18小0.068 鸥科 Laridae 红嘴鸥 Larus ridibundus40中7.017 红嘴巨鸥 Sterna caspia47~54大0.003 黑嘴鸥 Larus saundersi32中0.003 灰林银鸥 Larus heuglini60大0.033
说明: 体长数据引自文献[28], 种群占比数据来自《监测报告(2017年度)》。
Evaluation and Promotion Countermeasures of Bird Carrying Capacity of Mangrove Wetland in Guangdong-Hong Kong-Macau Greater Bay Area: A Case Study of Futian Mangrove, Shenzhen Bay
Abstract Focus on bird carrying capacity assessment in wetland ecological health assessment, the conversion method of ash free dry weight (AFDW) data of benthos was established, and the classification standard was optimized based on bird length. Taking Futian mangrove wetland as an example, the total food mass of benthos, the maximum calories for shorebirds and the field metabolic rate of shorebirds population were calculated. Furthermore, the carrying capacity of shorebirds was evaluated. The results are as follows. 1) The total food mass of benthos in Futian mangrove wetland had evident seasonal characteristics: winter (4.67×104kg) < spring (6.08×104 kg)<summer(8.00×104 kg)<autumn(1.23×105 kg). In autumn, the food mass density (in AFDW) of different habitats was as follows: mangrove area (89.22 g/m2) > beach area (3.58 g/m2) > gei wei fish pond area (0.22 g/m2). 2) The maximum calories for shorebirds in different seasons were winter (1.03×108 kJ) < spring (1.36×108 kJ) < summer (1.76×108 kJ) < autumn (2.70×108 kJ). 3) The field metabolic rate of the shorebirds population in Futian mangrove wetland was 467.27 kJ/d. 4) Autumn, winter and spring were the migration periods for migratory birds, and the carrying capacity of Futian mangrove wetland to shorebirds was 6431, 2438 and 3235 birds respectively. The actual observation data was higher than the research results, indicating that the current carrying capacity of shorebirds in the Futian mangrove wetland was insufficient to meet the food needs of birds in migration season, and the number of shorebirds might be reduced. It is suggested that the ecological protection of mangrove wetland in Futian should be strengthened in the following three aspects: the restoration and reconstruction of mangrove vegetation, the ecological restoration and functional improvement of gei wei fish pond, and the dynamic monitoring of birds and benthos. In this way, the total food mass of benthos and the bird carrying capacity can be lifted.
Key words Shenzhen Bay; mangrove wetland; benthos food mass; bird carrying capacity
doi: 10.13209/j.0479-8023.2020.066
广东省海洋经济发展专项([2020]059号)和深圳市科创委自然科学基金重点项目(JCYJ20200109140605948)资助
收稿日期: 2019-10-24;
修回日期: 2019-12-25