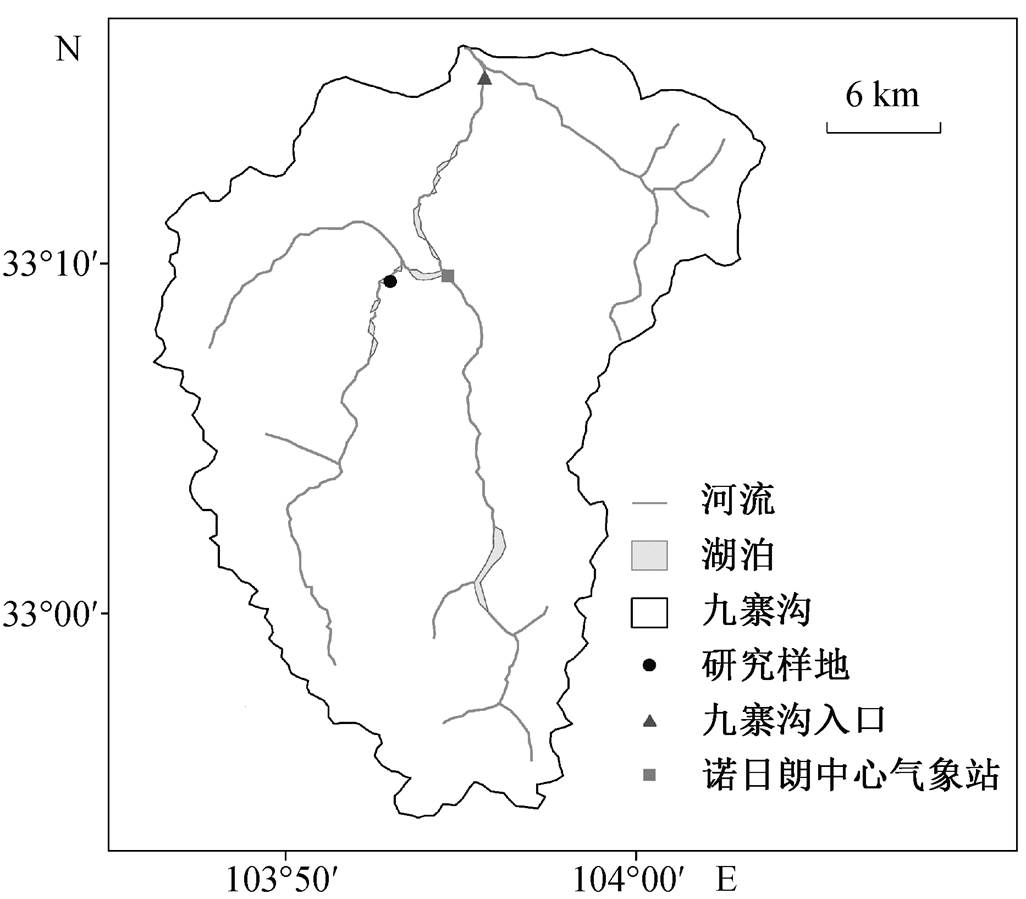
图1 九寨沟研究区域示意图
Fig. 1 Location of Jiuzhaigou Valley
摘要 利用 Granier 热扩散探针测定九寨沟针阔混交林 3 个主要树种 2013 年的树干液流密度, 分析各树种夜间液流的季节变化特征及夜间茎干补水与夜间蒸腾的分配特征。结果表明, 红桦(Betula albosinensis Burk.)和茶条槭(Acer ginnala Maxim.)的夜间液流占全天液流比例多为 0~30%, 油松(Pinus tabulaeformis Carr.)多为 0~25%, 但在某些特定条件下, 夜间液流占比会超过 40%; 生长季各树种夜间液流占比均呈“U”型变化, 生长季初期和末期较高, 生长季中期较低; 典型晴天内, 各树种夜间液流密度均与饱和水汽压差呈极显著的线性关系(R2>0.95), 表明夜间茎干补水和夜间蒸腾同时发生; 红桦、茶条槭和油松的夜间茎干补水和蒸腾分别占夜间液流活动的 80.7%和 19.3%, 81.4%和 18.6%, 63.9%和 36.1%。
关键词 夜间液流; 夜间茎干补水; 夜间蒸腾; 热扩散探针; 针阔混交林; 九寨沟
植物夜间液流指夜间土壤水分进入根系后, 通过输导组织运送至植物冠层, 最终存储在植物茎干(即夜间茎干补水)或者通过气孔蒸腾到大气中(即夜间蒸腾)的过程。由于过去的液流技术难以准确地测低液流速率, 微气象技术无法准确地测量夜间湍流数据[1], 因此之前的研究大多认为夜间植物气孔处于关闭状态[2], 植物不存在夜间液流现象。近年来, 随着液流技术和微气象技术的发展, 对不同物种、不同生态系统和不同生境条件下的野外观测表明, 植物夜间液流占全天液流的5%~25%[3]。
植物的夜间茎干补水(缓解日间木质部水势减小带来的压力)与夜间蒸腾(实际水分损失)可能同时或先后发生, 目前没有统一的技术进行定量的分割[4]。Fisher 等[1]采用 4 种方法分析同一树种的夜间液流分配特征, 结果中夜间茎干补水与夜间蒸腾量的占比因方法而异, 4 种方法的夜间茎干补水占比分别为 100%, 70%, 85%和 40%, 夜间蒸腾量占比分别为 0, 30%, 15%和 60%。夜间蒸腾量的不确定性会给生态系统水分收支研究结果带来一定的误差。在全球气候变化的背景下, 夜间饱和水汽压差和温度等环境条件会持续发生变化, 从而影响夜间液流过程及其在茎干补水和夜间蒸腾的分配特征, 使得夜间蒸腾对单木乃至生态系统的水量平衡影响的不确定性增加。因此, 需要对夜间液流组成进行准确的定量研究。
理论研究[5]和实测结果[6–7]均表明, 夜间饱和水汽压差是引起夜间蒸腾的主要因素。当夜间饱和水汽压差很小或为零时, 夜间液流主要用于茎干补水;当夜间饱和水汽压差较高, 且夜间液流与饱和水汽压差表现出显著的线性关系时, 夜间液流则主要用于植物的夜间蒸腾。本文利用 Granier 热扩散探针, 对九寨沟针阔混交林主要树种油松(P. tabulaeformis Carr.)、红桦(Betula albosinensis Burk.)和茶条槭(Acer ginnala Maxim.)进行观测, 研究不同季节夜间液流特征及其在夜间茎干补水和夜间蒸腾之间的分配特征, 旨在提高森林蒸腾和水量平衡估算的精确度, 为当地水资源管理和决策提供科学依据。
实验样地位于九寨沟国家自然保护区内北京大学森林生态系统地面定位观测研究站的针阔混交林(图 1)。九寨沟国家自然保护区(32°53′—33°20′N, 103°46′—104°05′E) 地处四川省阿坝藏族羌族自治州九寨沟县, 境内南北长 40.5km, 东西宽 35.4km[8], 全区面积为 720km2, 外围(核心景区以外的保护区)面积为 600km2, 流域面积(九寨沟汇水面积)为 651km2[9–10]。九寨沟处于中国北亚热带秦巴湿润区与青藏高原波密–川西湿润区的过渡地带, 在高原季风和亚热带海洋季风的影响下, 主要表现为冷干的季风气候特征。年均气温为 7.3℃, 最冷月(1 月)和最热月(7 月)的气温分别为–8.7℃和 16.8℃。年平均相对湿度为 60%~70%, 年平均降水量为 762mm, 且年变率较小(10%~15%)[11]。研究区干、湿季明显, 降水集中在 4—10 月, 占全年降水量的 70%以上。
图1 九寨沟研究区域示意图
Fig. 1 Location of Jiuzhaigou Valley
九寨沟属于中国西南地区典型的深切割高山峡谷地貌, 地势总体上南高北低, 海拔从 2000m (九寨沟沟口)到 4764m (尕尔纳峰)。受南北高差影响, 九寨沟气候垂直变化特征显著, 植物类型以及组成也呈现显著的垂直分布特征, 从高到低分别是针阔混交林带、亚高山针叶林带以及灌丛草甸带。针阔混交林带分布在海拔 1800~2800m, 主要包括黄果冷杉(Abies recurvata Mast. var. ernestii (Rehder) C. T. Kuan)和青杄(Picea wilsonii Mast.)等天然针叶树以及自然或人为破坏后更新的针叶树种(如油松); 还包括天然或人为更新的次生阔叶林, 主要有山杨(Populus davidiana Dode)、白桦(Betula platyphyllaSuk.)、红桦和辽东栎(Quercus liaotungensis Koidz.)等; 林内还有散生的茶条槭等阔叶树。此外, 很多原有的天然阔叶林内已有针叶树更新苗林, 从而形成针阔混交林。
在文献调研和实地勘察的基础上, 选取遭到破坏后人为更新的以油松为主的针阔混交林实验场地(33°9.54′N, 103°52.86′E; 海拔为 2478m), 选择油松、红桦和茶条槭为研究对象, 分别安装 Granier热扩散探针(thermal dissipation probe, TDP)[12–13]进行树干液流连续监测。其中, 油松为更新后主要树种, 红桦为主要次生林树种, 茶条槭散生于该混交林内。样树的基本特征见表 1。本研究重点比较油松、红桦和茶条槭的夜间树干液流和分配特征。
分别在各树种南北两个方位树高 1.5m 处刮去树干粗皮, 用特定规格的钻头在竖直方向分别横切向垂直于树心钻取直径为 1.2mm, 深度为 30mm 的两个孔洞, 孔洞间竖直距离为 15cm。分别插入 SF-G 型 Granier 热扩散探针(Ecomatic Ltd., 德国)。利用 CR1000 数据采集器(Campbell Scientific, 美国)采集树干液流速, 每分钟采集 1 次数据, 并存储每10 分钟的平均值。
在研究样地高约 30m 的高塔上(高于树冠 10m左右)分别安装 SKH2060 温湿度传感器(Skye, Inc., 英国)以及 CNR-4 四分量辐射传感器(Kipp & Zonen, 荷兰), 用于测量气温、相对湿度和辐射条件。通过 CR3000 数据采集器(Campbell Scientific, 美国)采集存储数据, 原始数据采集频率为 10Hz, 并记录每 10 分钟气温、相对湿度和辐射通量等数据的平均值。
本研究利用 Granier 热扩散探针, 直接测得树干上下两个探针之间的瞬时温度差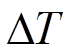 (°C), 再根据Granier[13]提出的树干液流密度(sap flux density)经验公式, 求得瞬时树干液流密度
(°C), 再根据Granier[13]提出的树干液流密度(sap flux density)经验公式, 求得瞬时树干液流密度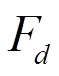 (g/(m2·s)):
(g/(m2·s)):
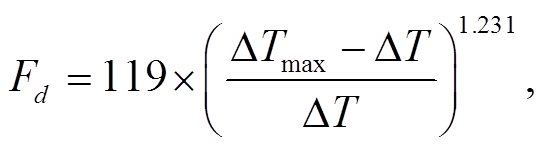
 (1)
(1)其中, 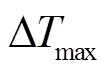 (°C)为昼夜最大瞬时温度差。一般而言, 温差最大时, 夜间树干液流密度为零。研究表明, 夜间零液流密度只有在饱和水汽压差 VPD 较小时才会出现[14]。
(°C)为昼夜最大瞬时温度差。一般而言, 温差最大时, 夜间树干液流密度为零。研究表明, 夜间零液流密度只有在饱和水汽压差 VPD 较小时才会出现[14]。
本研究采用美国杜克大学 Ram Oren 碳水生态实验室(C-H2O Ecology Lab Group)开发的 BaseLiner 3.0.11, 根据 10 分钟气象数据, 分别计算得到饱和水汽压差和净辐射通量。将净辐射通量小于零(夜间), 且连续两个小时内饱和水汽压差小于 0.05kPa时的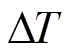 取为, 即此时无树干液流(Fd=0)。然而由于
取为, 即此时无树干液流(Fd=0)。然而由于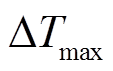 不恒定, 会随环境温度的变化而变化, 因此 BaseLiner 需先识别零液流密度条件来确定, 然后通过相邻线性插值来确定基线, 最后根据式(1)得到瞬时树干液流密度。为了减小测量方位引起的误差, 本研究采用南北两侧的树干液流密度平均值进行数据分析。
不恒定, 会随环境温度的变化而变化, 因此 BaseLiner 需先识别零液流密度条件来确定, 然后通过相邻线性插值来确定基线, 最后根据式(1)得到瞬时树干液流密度。为了减小测量方位引起的误差, 本研究采用南北两侧的树干液流密度平均值进行数据分析。
表1 研究区样树的基本特征
Table 1 Characteristics of three main species
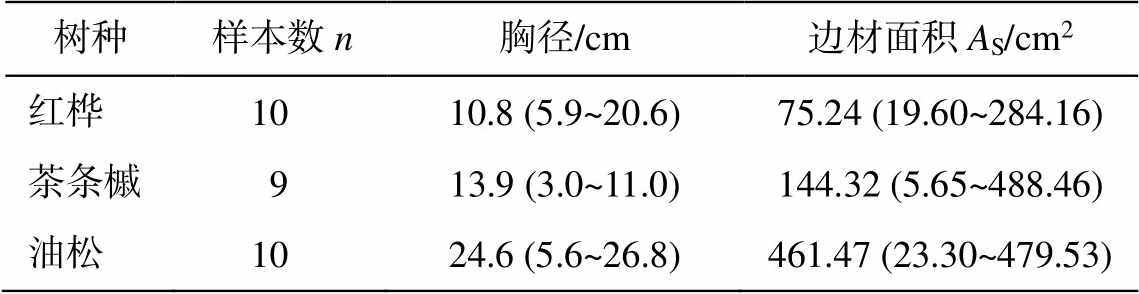
树种样本数n胸径/cm边材面积AS/cm2 红桦10 10.8 (5.9~20.6)75.24 (19.60~284.16) 茶条槭9 13.9 (3.0~11.0)144.32 (5.65~488.46) 油松10 24.6 (5.6~26.8)461.47 (23.30~479.53)
说明: 括号中数据分别为样树胸径(DBH)和样树胸径高度边材面积的范围。本研究利用 Haglof 树木生长锥钻取树心, 并通过染色法确定边材面积。
饱和水汽压差根据式(2)[15]进行计算:
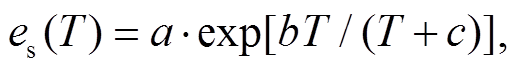 (2)
(2)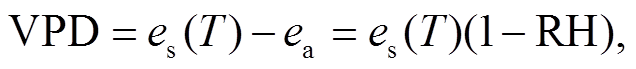 (3)
(3)
其中, 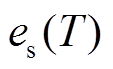 是 T 温度下的饱和水汽压(kPa); ea 是 T温度下空气中实际水汽压(kPa); a, b 和 c 为参数, 取值分别为 0.611kPa, 17.502, 240.97℃, VPD 为饱和水汽压差(kPa)。
是 T 温度下的饱和水汽压(kPa); ea 是 T温度下空气中实际水汽压(kPa); a, b 和 c 为参数, 取值分别为 0.611kPa, 17.502, 240.97℃, VPD 为饱和水汽压差(kPa)。
考虑到不同天气状况(晴天、阴天或多云天、雨天)下不同树种树干液流的日变化特征差异, 分别选取 2013 年 8 月 15 日、9 月 26 日和 8 月 28 日作为典型晴天、阴天和雨天。晴朗天气下, 各种典型树种树干液流密度曲线均为单宽峰型(图 2)。在树干液流启动前, 各树种树干液流密度趋近 0。随后, 树干液流密度随太阳辐射的增强而不断增加, 并与太阳辐射在 12:00 左右同时达到峰值。12:00—16:30, 虽然太阳辐射逐渐减弱, 但由于同期饱和水汽压差的持续增加, 各树种树干液流均维持在较高水平。16:30 以后, 太阳辐射量和饱和水汽压差同时减小, 各树种树干液流密度逐渐降低, 直到第二天 00:00 左右再次趋近于 0。在阴天条件下, 由于云量变化会引起太阳辐射的波动, 所以各树种的树干液流日内变化波动较大(图 3)。在典型降雨天气条件下(当日降水量为 22.8mm), 各树种树干液流密度显著低于晴天和阴天, 且红桦和茶条槭日内峰值出现在当日太阳辐射量和饱和水汽压差均较大的时段(图 4)。总体来看, 树干液流日变化主要受太阳辐射和饱和水汽压差的控制[16]。
由图 2~4 可知, 不同树种的树干液流速率有显著的差异, 各树种的日最大树干液流密度和日均树干液流密度排序为茶条槭>红桦>油松。受净辐射和饱和水汽压差的共同影响, 各树种平均树干液流密度排序为晴天>阴天>雨天。各典型天气情况下, 各树种均表现出明显的昼夜变化规律, 夜间(18:00后)存在明显的液流活动。
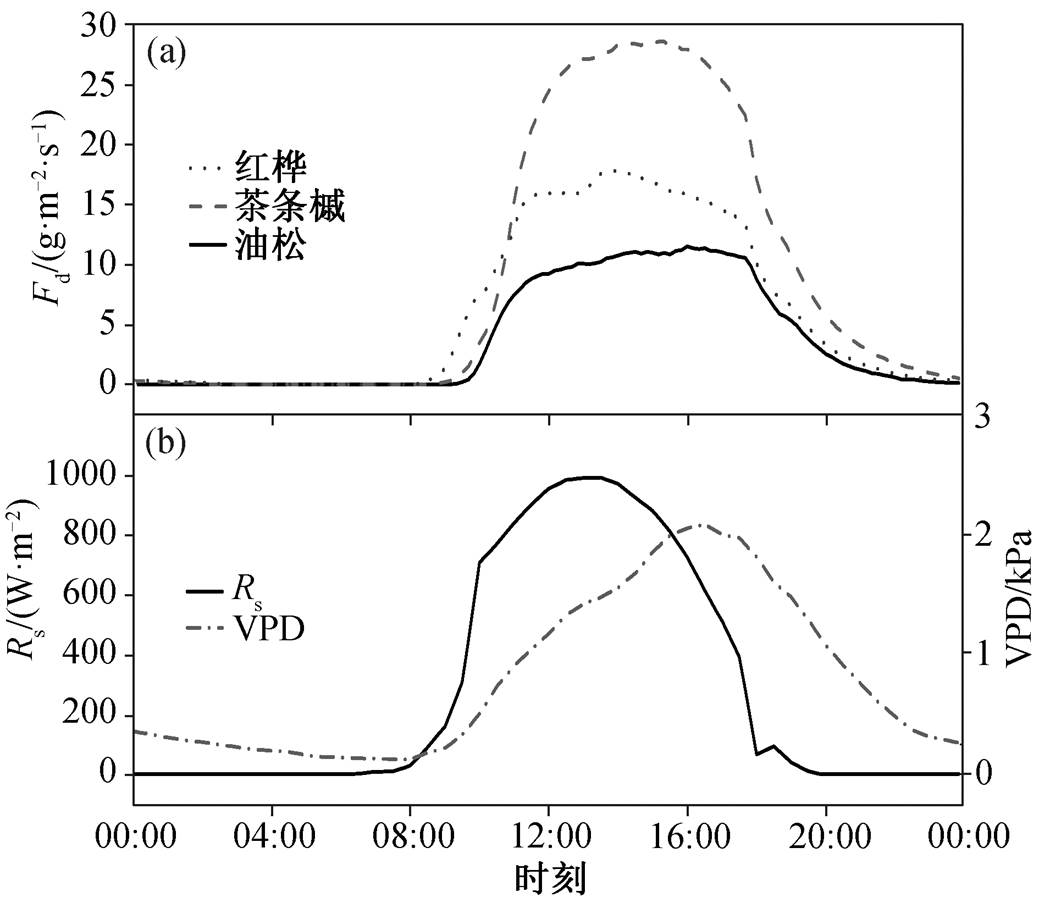
图2 2013 年生长季典型晴天(8 月 15 日)不同树种的树干液流密度、太阳辐射(Rs)和饱和水汽压差的日内变化
Fig. 2 Diurnal course of sap flux density for dominant species in a typical sunny day (August 15, 2013)
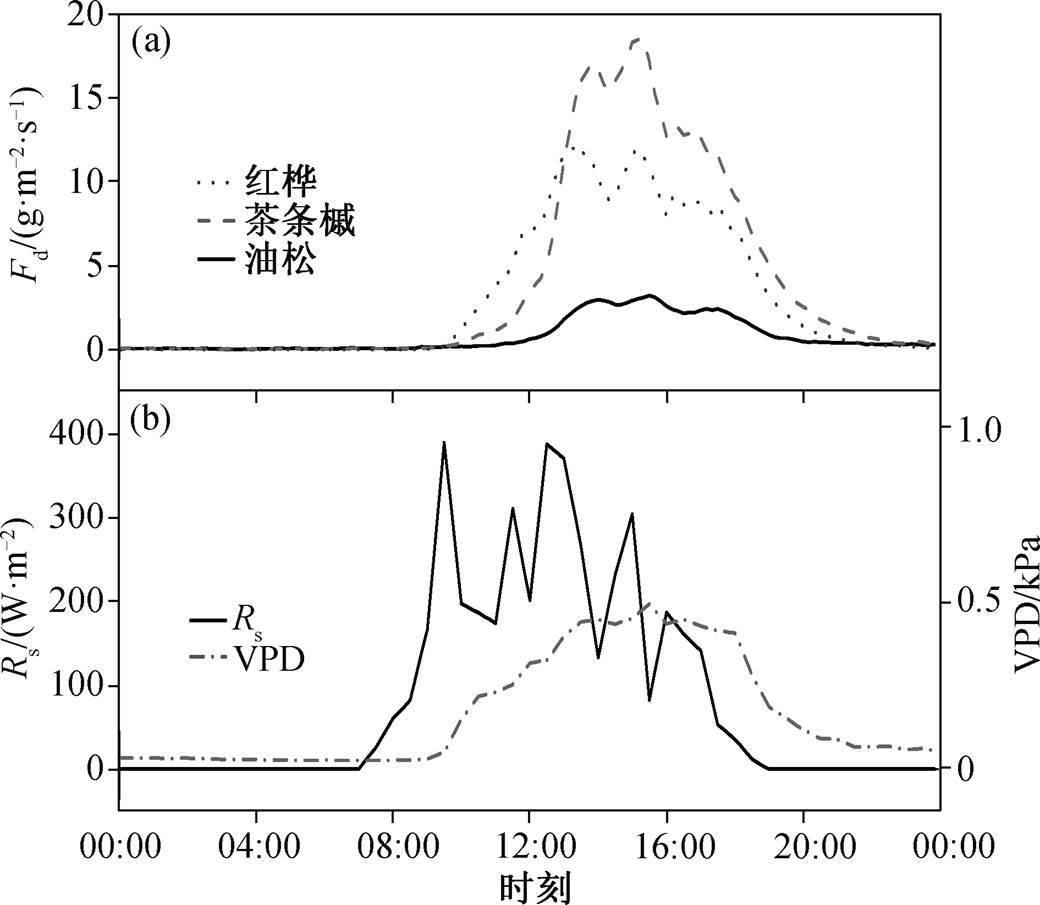
图3 2013 年生长季典型阴天(9 月 26 日)不同树种的树干液流密度、太阳辐射和饱和水汽压差的日内变化
Fig. 3 Diurnal course of sap flux density for dominant species in a typical cloudy day (September 26, 2013)
研究区针阔混交林中红桦、茶条槭和油松液流速率季节变化特征如图 5 所示。可以看出, 2013 年红桦树干液流启动时间为 4 月 30 日, 茶条槭为 4 月24 日, 均明显晚于油松, 可能是由于研究区阔叶树新叶主要在 4 月下旬开始生长。与红桦和茶条槭相比, 油松液流可以维持更长的时间, 结束时间(11月 13 日)也明显晚于红桦和茶条槭(10 月 23 日)。从液流速率看, 红桦液流密度在 0~23g/(m2·s)之间, 茶条槭液流密度在 0~36g/(m2·s)之间, 油松在 0~15g/(m2·s)之间。孙鹏森等[17]对华北地区主要造林树种油松的液流速率观测结果表明, 油松日最大液流密度为 20g/(m2·s)左右。温杰等[18]发现黄土丘陵区人工林中油松日最大液流密度可高达 60g/(m2·s)。本研究区油松为森林砍伐后自然演替树种, 液流密度更低。
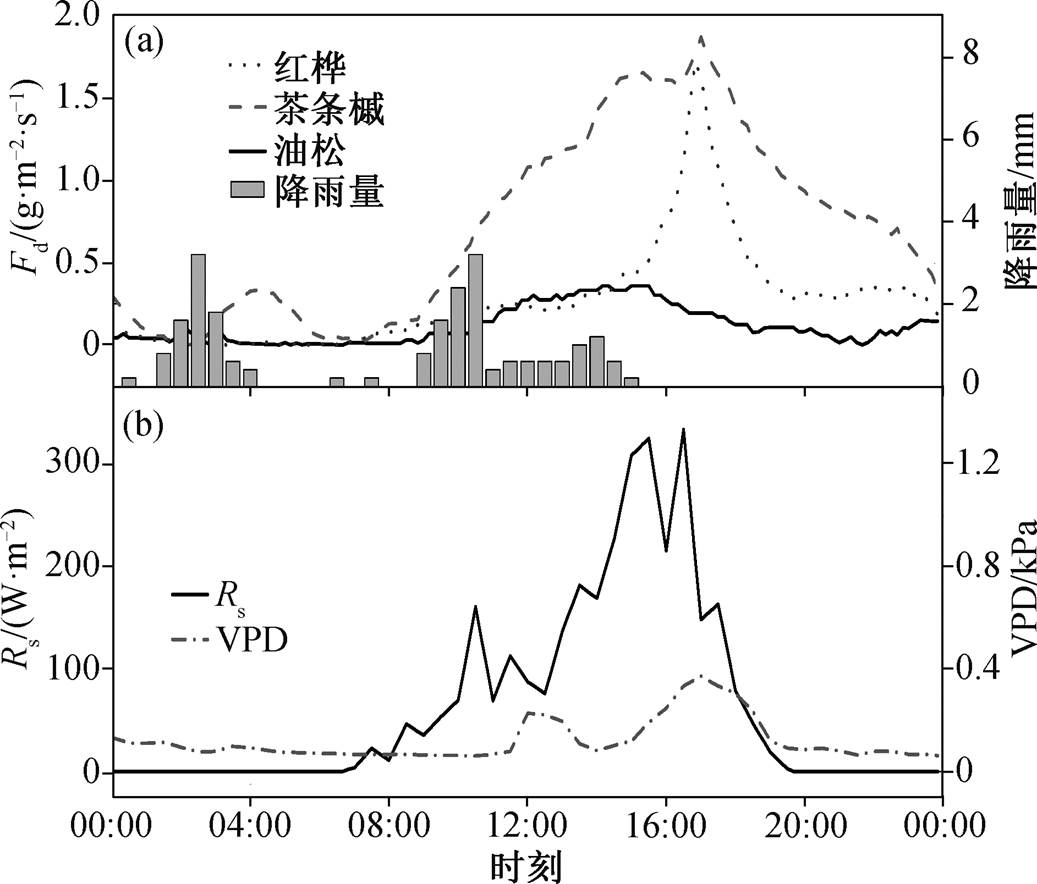
图4 2013 年生长季典型雨天(8 月 28 日)不同树种的树干液流密度、太阳辐射和饱和水汽压差的日内变化
Fig. 4 Diurnal course of sap flux density for dominant species in a typical rainy day (August 28, 2013)
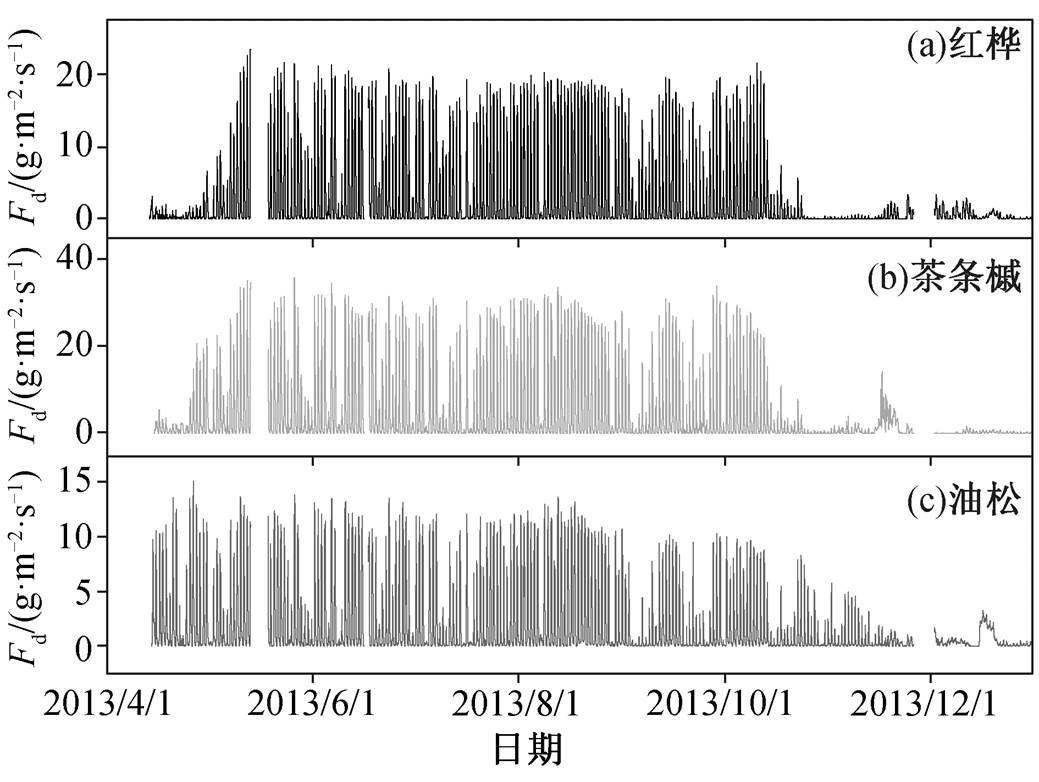
图5 研究区 2013 年 4—12 月红桦、茶条槭和油松树干液流密度季节变化特征
Fig. 5 Seasonal variation of sap flux density for Betula albosinensis Burk., Acer ginnala Maxim., and Pinus tabulaeformis Carr. from April 14 to Decem-ber 31 in 2013
在已有的研究中, 夜间液流的大小通常用占白天液流或全天液流的比例来表示[3]。为了进一步比较不同树种夜间液流的差异, 本研究对 2013 年生长季各树种夜间液流占全天液流的比例以及该比例的分布频数进行分析, 发现生长季中各树种夜间液流/全天液流均呈“U”型变化, 即生长季初期(4 月底)夜间液流占比较大, 随后(5 月)逐渐减小, 并在生长季中期(6—8 月)维持较高的水平, 最后在 9—10 月逐渐增大(图 6(a)、6(c)和 6(e)), 与尹立河等[19]对陕西榆林地区旱柳和小叶杨夜间树干液流季节变化特征的研究结果一致。
不同树种夜间液流的大小存在一定程度的差异。红桦夜间液流占全天液流的比例在 0~55.6%之间, 全生长季平均值为 12.5%; 茶条槭夜间液流占全天液流的比例在 0~68.5%之间, 全生长季平均值为 14.9%; 油松夜间液流占全天液流的比例在 0~ 43.5%之间, 全生长季平均值为 12.9%。从夜间液流大小频数分布(图 6(b)、(d)和(f))可知, 红桦夜间液流占全天液流的比例多在 0~30%之间(该范围内天数/生长季天数=175/186, 下同), 茶条槭多在0~30%之间(175/186), 油松多在 0~25%之间(176/ 186)。但是, 在某些特定条件下, 夜间液流的占比超过 40%, 与相关实测结论[7]一致。
一般认为, 夜间液流用于夜间蒸腾和补给树干储水量[1,14]。当夜间液流多用于夜间蒸腾时, 饱和水汽压差 VPD 通常是一个关键的推动力[5–7,20–22]。若夜间 VPD 为 0, 则对夜间蒸腾无影响。本研究区在高原季风和亚热带海洋季风的影响下, 主要表现为冷干的季风气候特征。如图 7 所示, 典型晴天夜间(19:50 至次日凌晨 6:00) VPD 多在 0.2kPa 以上, 且各树种夜间液流密度均与 VPD 表现出极显著的线性关系(红桦: R2 = 0.98, p<0.001; 茶条槭: R2=0.99,p<0.001; 油松: R2=0.95, p<0.001)。这说明典型晴天条件下, VPD 是该地区夜间蒸腾的驱动力, 各树种在进行夜间储水量补给的同时进行夜间蒸腾。
为了研究夜间液流的分配特征, 本文以 2013年 8 月 15 日为例, 估算红桦、茶条槭和油松的夜间茎干补水量和夜间蒸腾量。首先, 对该典型睛天各树种树干液流密度与太阳辐射进行交叉相关分析(cross correlation analysis), 发现各树种树干液流密度的日内变化均滞后太阳辐射 80 分钟。在滞后期间 (19:50—21:10), 夜间液流活动因缺乏持续的动力(太阳辐射)而无法通过植物气孔以蒸腾的形式扩散出去, 因此夜间液流基本上用于茎干补水活动(图 8 中黑色阴影区)。滞后期之后(21:10 后), 由于VPD 仍然较高, 且与液流密度极显著正相关, 夜间茎干补水和夜间蒸腾同时进行, 夜间茎干补水速率开始逐渐衰减, 衰减速率为滞后期结束时刻(21:10)曲线的斜率, 树干液流密度曲线、滞后期结束时刻曲线的切线以及横坐标围绕区域的面积则为夜间较高 VPD 条件导致的夜间蒸腾量(图 8 中灰色阴影区)。尽管该典型睛天内红桦、茶条槭以及油松夜间液流占比(分别为 4.0%, 4.7%和 6.5%)差别不大, 但各树种夜间液流的分配不尽相同。根据各区域积分面积可知, 红桦夜间茎干补水和蒸腾分别占夜间液流活动的 80.7%和 19.3%, 茶条槭分别占 81.4%和18.6%, 油松分别占 63.9%和 36.1%。Fisher 等[1]基于类似的方法, 研究美国加利福尼亚 Tonzi Ranch试验区蓝栎的夜间液流活动, 发现夜间蒸腾水分损失占夜间液流活动的比例与土壤水分条件有关, 在干旱区域, 夜间液流主要用于夜间茎干补水, 夜间蒸腾占比(10.5%)较小。本研究区土壤水分更为充足, 夜间蒸腾占比更高, 在估算植物蒸腾耗水时, 夜间蒸腾不容忽视[23]。
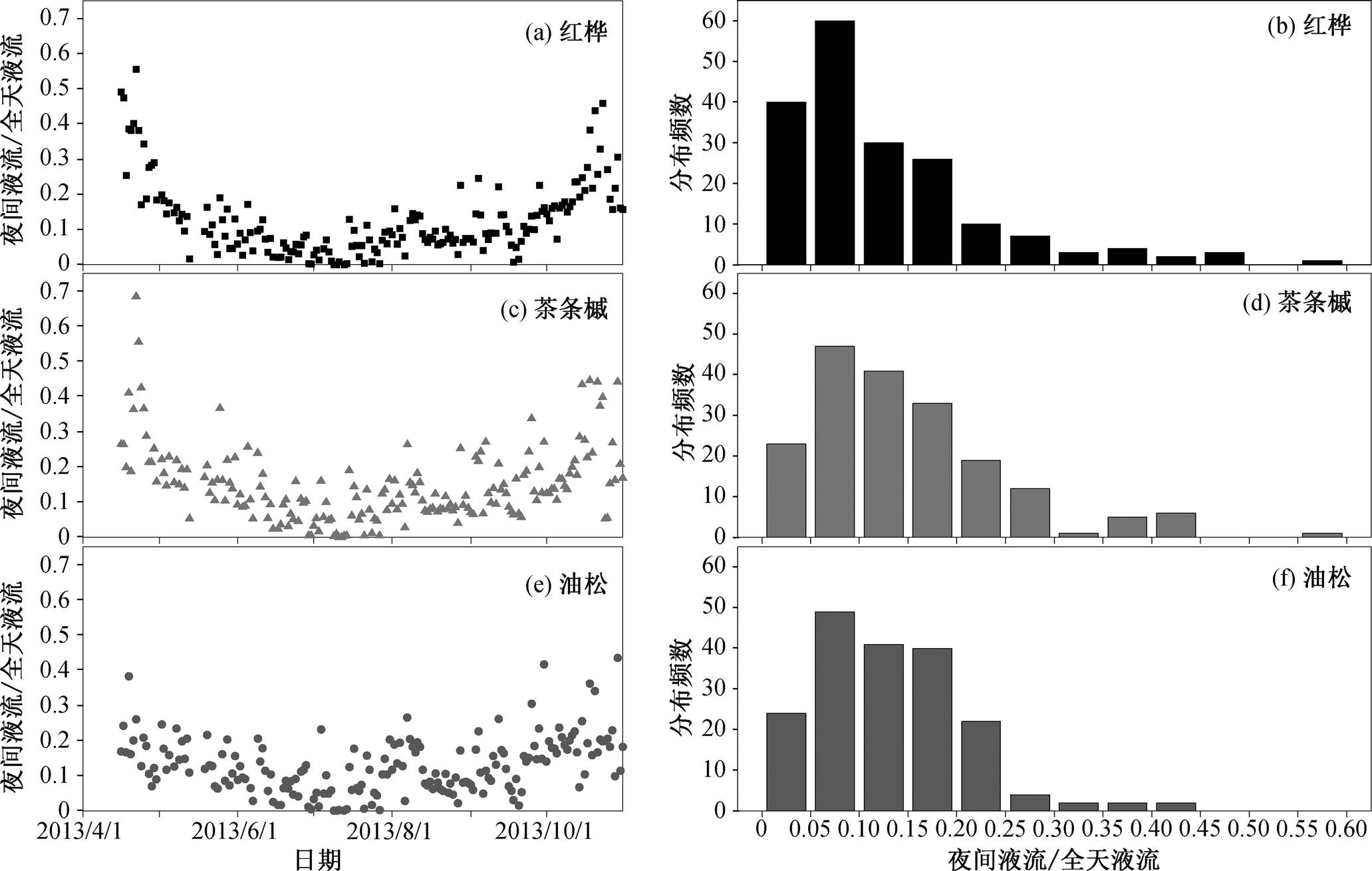
图6 研究区 2013 年生长季 4—10 月各树种的夜间液流占全天液流比例的季节变化特征和频数分布(n=186)
Fig. 6 Seasonal variation and frequency counts of the ratio of nighttime sap flow to daily rates during the growth season (from April to October) of 2013 for three main species (n=186)
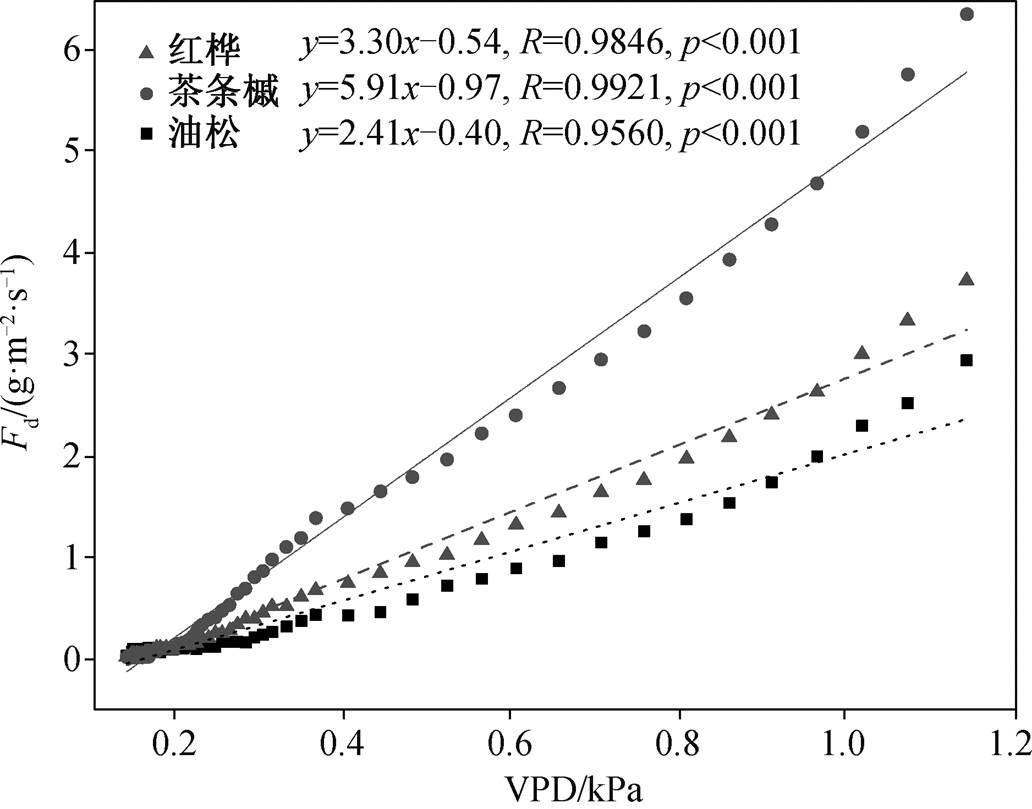
图7 典型晴天时各树种夜间液流密度与 VPD 的关系
Fig. 7 Results of linear fitting between nighttime sap flux density and vapor pressure deficit for dominant species in a typical sunny day
本研究基于九寨沟针阔混交林主要树种 2013年的树干液流观测数据, 分析红桦、茶条槭和油松夜间液流季节变化特征以及夜间茎干补水和夜间蒸腾的分配特征, 结论如下。
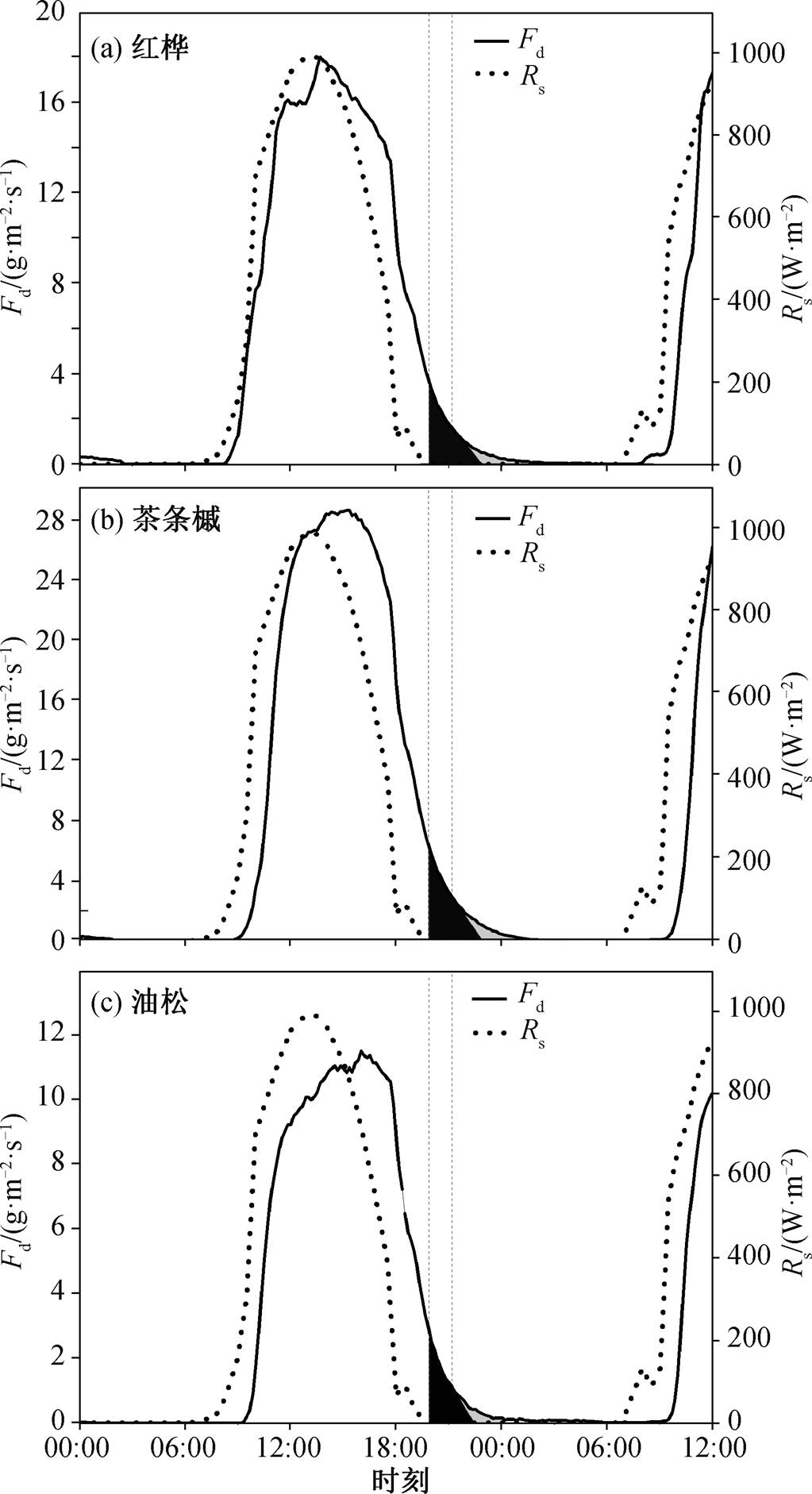
左侧灰色虚线代表太阳落山时间(19:50), 左右灰色虚线的间距表示液流活动与太阳辐射变化的滞后时间(80 分钟); 黑色阴影为夜间茎干补水, 灰色阴影为夜间蒸腾
图8 研究区生长季典型晴天(2013 年 8 月 15 日)各树种的夜间液流组成分配特征
Fig. 8 Estimation of nighttime transpiration and refilling for three main species in a typical sunny day ( August 15, 2013)
1)各树种平均树干液流密度均为晴天>阴天>雨天, 各树种树干液流密度排序为茶条槭>红桦>油松。
2)2013 年生长季, 各树种夜间液流占全天液流的比例均呈“U”型变化, 即生长季初期和末期较高, 中期较低。
3)在全生长季, 红桦、茶条槭和油松夜间液流占全天液流比例的平均值分别为 12.5%, 14.9%和12.9%。红桦夜间液流占全天液流的比例多在 0~ 30%之间, 茶条槭多在 0~30%之间, 油松多在 0~ 25%之间, 但在某些特定条件下, 夜间液流占比超过 40%。
4)典型晴天条件下, 各树种夜间液流密度均与VPD 表现出极显著的线性关系, 各树种夜间储水和夜间蒸腾同时进行。
5)红桦、茶条槭和油松夜间茎干补水与蒸腾分别占夜间液流活动的 80.7%和 19.3%、81.4%和18.6%、63.9%和36.1%。
致谢 九寨沟管理局科研处在场地和入沟许可等方面提供支持, 王振华工程师、柴民伟博士、硕士生向皎和王永强以及 Shinichi Takeuchi 教授等在仪器安装维护与数据分析等方面给予协助, 一并致以衷心感谢。
参考文献
[1] Fisher J B, Baldocchi D D, Misson L, et al. What the towers don’t see at night: nocturnal sap flow in trees and shrubs at two AmeriFlux sites in California. Tree Physiology, 2007, 27(4): 597–610
[2] Kramer P J, Boyer J S. Water relations of plants and soils. San Diego: Academic Press, 1995
[3] 方伟伟, 吕楠, 傅伯杰. 植物夜间液流的发生、生理意义及影响因素研究进展. 生态学报, 2018, 38(21): 7521–7529
[4] Huang C W, Domec J C, Ward E J, et al. The effect of plant water storage on water fluxes within the coupled soil-plant system. New Phytologist, 2017, 213(3): 1093–1106
[5] Monteith J L. Evaporation and environment. Sym-posia of the Society for Experimental Biology, 1965, 19(19): 205–234
[6] Zeppel M, Tissue D, Taylor D, et al. Rates of nocturnal transpiration in two evergreen temperate woodland species with differing water-use strategies. Tree Physiology, 2010, 30(8): 988–1000
[7] Dawson T E, Burgess S S O, Tu K P, et al. Nighttime transpiration in woody plants from contrasting ecosys-tems. Tree Physiology, 2007, 27(4): 561–575
[8] 林雯. 九寨沟自然保护区森林生态系统功能研究[D]. 成都: 四川大学, 2006: 28–34
[9] 邓贵平. 九寨沟旅游地学景观成因与保护. 北京: 科学出版社, 2012
[10] Hao Yunqing, Wang Jinxi, Jiang Hong. The dynamics of land cover change pattern and landscape fragmen-tation in Jiuzhaigou Nature Reserve, China. Procee-dings of SPIE—The International Society for Optical Engineering, 2009, 7498: 74983P1–17
[11] 杨俊义, 万新南, 范晓, 等. 九寨沟水环境系统动态模拟. 四川地质学报, 2002, 22(1): 16–20
[12] Granier A. A new method of sap flow measurement in tree stems. Annales Des Sciences Forestieres, 1985, 42(2): 193–200
[13] Granier A. Evaluation of transpiration in a Douglas-fir stand by means of sap flow measurements. Tree Physiology, 1987, 3(4): 309–320
[14] Oishi A C, Oren R, Stoy P C. Estimating components of forest evapotranspiration: a footprint approach for scaling sap flux measurements. Agricultural and Fo-rest Meteorology, 2008, 148(11): 1719–1732
[15] Campbell G S, Norman J M. An introduction to envi-ronmental biophysics. New York: Springer Science and Business Media, 2012
[16] 赵平, 饶兴权, 马玲, 等. Granier 树干液流测定系统在马占相思的水分利用研究中的应用. 热带亚热带植物学报, 2005, 13(6): 457–468
[17] 孙鹏森, 马履一, 王小平. 油松树干液流的时空变异性研究. 北京林业大学学报, 2000, 22(5): 1–6
[18] 温杰, 陈云明, 唐亚坤, 等. 黄土丘陵区油松、沙棘生长旺盛期树干液流密度特征及其影响因素. 应用生态学报, 2017, 28(3): 763–771
[19] 尹立河, 黄金廷, 王晓勇, 等. 陕西榆林地区旱柳和小叶杨夜间树干液流变化特征分析. 西北农林科技大学学报(自然科学版), 2013, 41(8): 85–90
[20] Benyon R G. Nighttime water use in an irrigated Eucalyptus grandis plantation. Tree Physiology, 1999, 19(13): 853–859
[21] Hogg E H, Hurdle P A. Sap flow in trembling aspen: implications for stomatal responses to vapor pressure deficit. Tree Physiology, 1997, 17(8/9): 501–509
[22] Oren R, Sperry J S, Ewers B E, et al. Sensitivity of mean canopy stomatal conductance to vapor pressure deficit in a flooded Taxodium distichum L. forest: hydraulic and non-hydraulic effects. Oecologia, 2001, 126(1): 21–29
[23] 王艳兵, 德永军, 熊伟, 等. 华北落叶松夜间树干液流特征及生长季补水格局. 生态学报, 2013, 33 (5): 1375–1385
Characteristics of Nighttime Sap Flow and Its Partition in a Mixed Forest in Jiuzhaigou Valley
Abstract To investigate the characteristics of nighttime sap flow and its partition into nocturnal water refilling and transpiration, the Granier-type thermal dissipation probes were used to measure sap flux density in 2013 in three main species of a mixed forest in Jiuzhaigou Valley. The results showed that the ratio of nighttime sap flow to daily value mainly ranged from 0 to 30% for Betula albosinensis Burk. andAcer ginnala Maxim., from 0 to 25% for Pinus tabulaeformis Carr., and might excess 40% in some special occasions. It showed a “U-shaped” seasonal variation, with lower ratios in the mid-growing season and higher ratios in the early and late growing season. There was a significant linear relationship between nighttime sap flow and vapor pressure deficit during a clear day, indicating the occurrence of nocturnal transpiration along with refilling. The ratios of nocturnal water refilling and transpiration to the total nighttime sap flow were 80.7% and 19.3%, 81.4% and 18.6%, 63.9% and 36.1%, respectively for the three tree species.
Keywords nighttime sap flow; nocturnal water refilling; nocturnal transpiration; thermal dissipation probe; mixed forest; Jiuzhaigou Valley
doi: 10.13209/j.0479-8023.2020.042
深圳市基础研究项目(JCYJ20180504165440088)资助
收稿日期: 2019–07–01;
修回日期: 2019–08–29