围封对蒙古荒漠草原和高山草原植物群落组成及稳定性的影响
廖晗茹 Indree Tuvshintogtokh 郭通† 赵景学
北京大学城市与环境学院, 地表过程分析与模拟教育部重点实验室, 北京 100871; †通信作者, E-mail: tongg@pku.edu.cn
摘要 基于蒙古国两种典型草地生态系统类型(荒漠草原和高山草原), 采取围封和放牧两种处理方式, 对2013—2018 年这两种草地类型的群落盖度进行系统分析。按植物分类学法, 将群落内的所有物种划分为五大功能群——杂草类、蒿类、豆科、禾本科和莎草科, 采用 Gordon 稳定性方法评估围封对群落稳定性的影响。结果表明: 1)围封显著增加荒漠草原杂草类植物覆盖度, 减少蒿类和禾本科植物覆盖度, 年际间无明显的变化规律, 而对于高山草原, 围封对不同功能群盖度无显著影响; 2)围封增加荒漠草原的群落稳定性, 而对高山草原无明显影响; 3)群落稳定性与优势种盖度占群落的比例呈显著的正相关关系, 荒漠草原的优势种占比高于高山草原, 导致荒漠草原的群落稳定性更好。根据上述结果, 建议在对草地进行围封管理前, 应充分考虑当地的环境条件及围封对植物群落及生态系统的潜在影响, 同时辅以休牧、轮牧或季节性放牧等措施, 才能真正提高草原生态系统服务的使用价值。
关键词 年际变化; 群落结构; 功能群; Gordon稳定性; 植物覆盖度
草地约占地球陆地总面积(除格陵兰岛和南极洲外)的 40.15%[1], 是陆地生态系统的重要组成部分。草地生态系统具有调节气候、涵养水源、防风固沙和改良土壤等生态服务和功能。放牧是影响草地生态系统结构和功能稳定性的重要因素, 其影响方式包括家畜对草地物种的啃食、踩踏和排泄物的分解等[2]。放牧可以改变植物群落的结构, 进而改变其稳定性。通常, 适度的放牧有利于草地群落多样性和生产力的维持。然而, 当前大多数草地生态系统遭受长期放牧的威胁[3–5]。为了减缓持续放牧对草地生态系统的影响, 围栏禁牧措施被广泛应用于草地的恢复管理中[6–7]。为评估围封对群落组成和稳定性的影响, 通常分析植被特征(如盖度和植被类型)的变动状况, 试验证据显示围封通常会增加草地群落的盖度[8–9]。为了更好地量化群落组成对围封的响应, 德米特里耶夫[10]按植物分类学的科组, 划分出禾本科、豆科、杂草类和莎草科四大功能类群。动物对不同科组喜食程度的差异会改变群落的组成, 研究发现禾本科类群比杂草类群更易被食草者摄食[11–12]。群落组成的变化会影响其结构的稳定性。主要原因是围栏封育改变了不同植物功能群在群落中的比例, 同时这种影响会受到区域气候的调控[13–14]。研究表明, 由于资源可获取性的差异, 草地中的豆类和禾本科对群落稳定性的影响是不同的[15]; 青藏高原高寒草地试验表明, 围封显著增加多年生禾本科类群的丰富度, 却降低群落的稳定性[16]。
可以从结构的变动性、植物功能的变动性、群落对环境压力的抵抗能力、群落的自组织能力和恢复能力等多方面评估群落稳定性[17–18]。找到适用于评估草地群落稳定性的方法和标准, 对于草地的管理和恢复具有重要的理论和现实意义。Gordon稳定性操作方法简单, 标准统一。该稳定性概念源于工业领域, 生态学家将其引入植物群落研究中, 主要用于群落结构的评估, 即假定物种数目和频度按照某种排列组合时, 群落结构达到稳定状态, 群落内物种不易消失, 外来物种不易入侵[19]。物种消失易导致群落某种植物功能的丧失, 而入侵物种会改变群落的原有组成, 二者均会对群落稳定性产生影响。Gordon 稳定性本质上由群落中的优势种决定, 优势种通常主导植物群落功能的变化[20]。群落中优势种比重与Gordon 稳定性的定量关系在很多研究中被忽视。Gordon 稳定性方法仅能判别群落结构稳定与否, 无法提供群落演替的信息。此外, 可以将 Gordon 稳定性方法近似地视为一种经验模型, 其缺点是没有考虑群落结构变化的机理, 这也表明该方法的局限性。研究发现, 围封导致青海湖流域高寒草甸群落 Gordon 稳定性的降低, 但促进高寒草原群落稳定性的增加[14]。围封年限也会对群落 Gordon 稳定性产生影响, 短期围封有助于围栏内群落 Gordon 稳定性的增加, 随着围封时间的增加, 围栏外的群落显得更加稳定[16]。
为了深入研究草地生态系统群落组成与稳定性之间的关系, 我们选取蒙古国草地作为研究对象。草地占蒙古国土面积的 83.4%[21], 游牧自古以来为该国最主要的人类活动, 因此研究围栏封育对草原生态系统群落稳定性的影响十分具有代表性。前人仅对蒙古围封草地植被特征及多样性等进行单维度调查研究[21–22], 对其群落稳定性的研究相对匮乏。本文以高山草原和荒漠草原为研究对象, 通过围封试验, 并结合长期定位调查的群落物种类型和盖度等数据, 探讨蒙古国典型草地群落组成和稳定性对围栏封育的响应, 以期为地区退化草地的恢复和草地植被的管理提供科学依据。
1 材料与方法
1.1 试验区域概况
试验地点(47°112.838′N, 108°144.240′E)位于蒙古高原, 海拔 1235m, 距离蒙古国首都乌兰巴托约250km。冬季(11 月至次年 4 月下旬)气候干冷, 夏季(6 月至 9 月)温暖多雨, 生长季通常为 4 月下旬至10 月下旬。试验区气候、土壤条件及植物群落特征如表 1 所示。
1.2 实验设计
设置高山草原和荒漠草原两种草地类型, 每种类型设置围封改良和正常放牧两种处理方式, 围栏外以正常放牧作为对照处理。高山草原和荒漠草原自 2013 年起进行封育, 分别在两种草地类型中随机选取 10 个面积为 1m×1m 的样地, 对各个样地进行长期的定位调查。2014 年后, 改为在荒漠草原中调查 6 个样地, 在高山草原中调查 3 个样地, 每年 7—8 月调查植物物种类型和盖度等指标。同时, 参考德米特里耶夫植物分类方法[10], 将两种草地类型的植物物种分为杂草类、蒿类、豆科、禾本科和莎草科 5 种功能群。杂草类主要包括百合科砂韭 Allium bidentatum、蔷薇科的星毛委陵菜 Potentilla acaulis和蕨麻 Potentilla anserine、环胚亚科的木地肤 Kochia prostrata和菊科的高山紫苑Aster alpinus等; 蒿类主要包括菊科蒿属植物, 如冷蒿 Artemisia frigida、东北丝裂蒿 Artemisia adamsii 和猪毛蒿 Artemisia scopari 等; 豆科包括乳白黄芪属的黄芪 Astragalus galacitites 和糙叶黄芪 Astragalus scaberrimus 以及野豌豆属的广布野豌豆 Vicia cracca L 等; 禾本科包括羊茅属的苔原羊矛 Festuca lenensis 和羊矛 Festuca ovina、针茅属狼针草 Stipa baicalensis 以及洽草属洽草 Koeleria macrantha 等; 莎草科包括薹草属的柄状薹草 Carex pediformis、寸草 Carex duriuscula和扁囊薹草Carex coriophora等。
表1 两种草地生态系统气候条件、土壤属性及群落特征
Table 1 Climate conditions, soil properties and community characteristics of two steppe ecosystems

项目荒漠草原高山草原 经纬度47°12N, 108°44E48°17'N, 108°43'E 海拔/m12521519 年均降水/mm258.4287.5 年均温/ºC−2.60−1.75 土壤类型棕钙土暗棕壤 植物覆盖度/%53.567.5 放牧强度/(bod·hm–2)0.08~0.140.04~0.09 优势种冷蒿Artemisia frigida砂韭Allium bidentatum东北丝裂蒿Artemisia adamsii木地肤Kochia prostrata乳白黄芪Astragalus galacitites星毛委陵菜Potentilla acaulis高山紫菀Aster alpinus苔原羊矛Festuca lenensis柄状薹草Carex pediformis狼针草Stipa baicalensis
说明: 由于 2018 年数据未更新, 数据为 2013—2017 年平均值, bod 为蒙古国计量单位, 1 bod 等价于 1 匹马、1 头牛、1 头牦牛或 6 只羊。
1.3 数据处理
1.3.1植物覆盖度
植物覆盖度的计算公式为
式中, n代表样地的个数, m代表某功能类群的物种数, Cij代表该功能类群第i个物种在第j个样地的盖度。
1.3.2 植物群落稳定性
本研究结合郑元润[23]和罗久富等[16]对 Gordon稳定性计算方法的改进。其中, 郑元润[23]的改进主要是通过建立数学方程确定交点坐标, 而不是利用在方格纸上手绘确定, 降低了人工操作时的误差。罗久富等[16]的改进体现为两点: 1) 将植物的频度替换为植物的盖度, 频度主要反映物种数量的信息, 盖度可以间接地反映群落功能(如生物量)的变化, 因此用植物覆盖度来度量群落结构的稳定性更具代表性[24]; 2)将一元二次方程改为一元三次方程进行拟合, 其目的是提高拟合的精度, 幂函数的指数越大, 拟合的精度通常会越高。罗久富等[16]改进后的 Gordon 稳定性计算方程如下:
其中, X为种累积百分数, i代表第i个物种, S为总物种数;
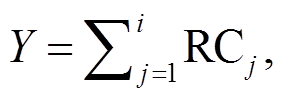 (3)
(3)
其中, Y代表累积相对盖度, RC代表相对盖度。
以种累积百分数为横轴 x, 累积相对盖度为纵轴 y, 将各物种对应的 x, y 作图, 用如下曲线方程拟合。
平滑曲线拟合方程为
y = ax3 + bx2 + cx + d , (4)
直线方程为
y = 100 – x , (5)
将式(5)带入式(4)得
ax3 + bx2 + (c+1) x + d −100 = 0。 (6)
解得 x 后, 根据实际情况, 选取x, y 均为实数且大于 0 的结果, 计算坐标点(x, y)与坐标点(20, 80)的欧氏距离, 该欧氏距离数值即可量化群落稳定性。很多研究表明, 当草地群落植物种总数倒数累积和累积相对频度(或盖度)满足坐标点(20, 80)时, 物种组成相对稳定, 即现有的物种组成基本上维持原有的物种组成[16,23,25]。
采用 R 统计软件, 对不同植物功能群围栏内外的盖度进行 T 检验, 量化围封对功能群组成的影响。利用 Origin2018 软件完成绘图。
2 结果
2.1 围封对植物群落组成的影响
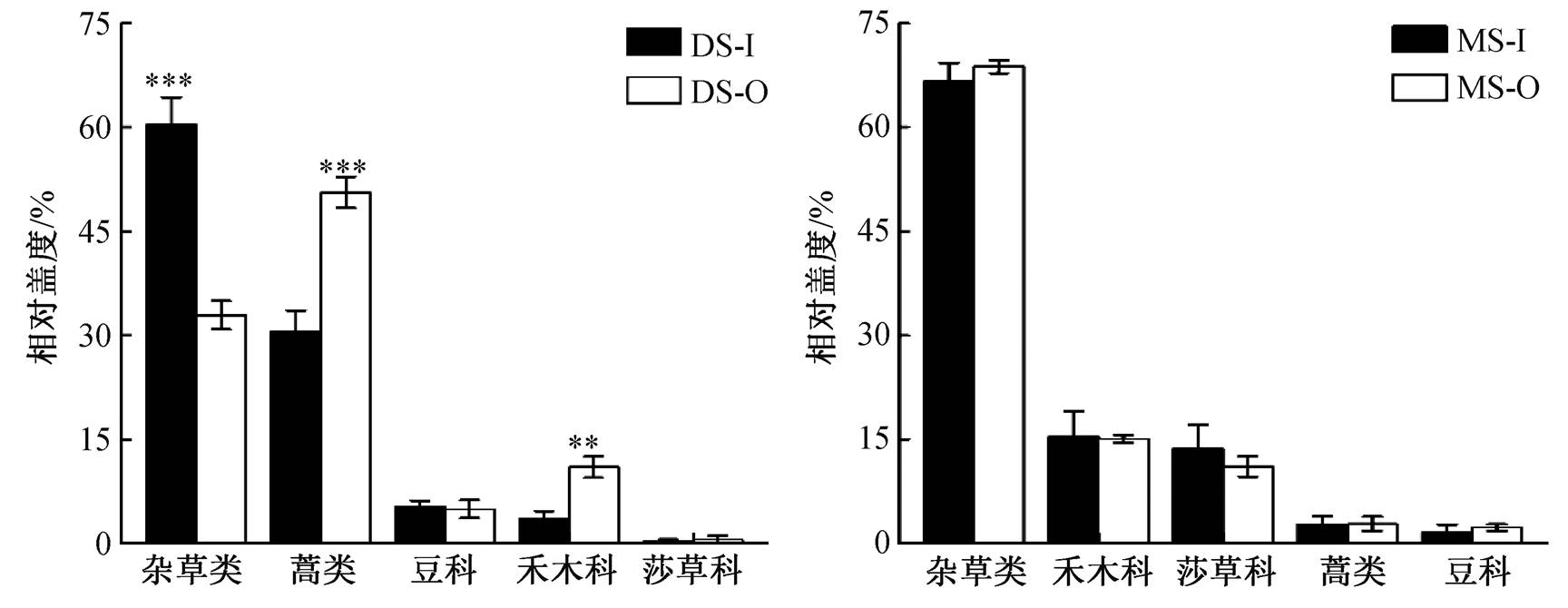
如图 1 所示, 围封显著提高荒漠草原杂草类功能群的覆盖度(P<0.001), 显著降低蒿类和禾本科功能群的覆盖度(P<0.01)。然而, 高山草原各功能群覆盖度对围封的响应并不显著。在荒漠草原中, 围栏外蒿类占比最高, 为 48.92%, 其次为杂草科, 占比32.03%, 后面依次为禾本科、豆科和莎草科, 占比分别为 10.72%, 5.53%和 2.80%。围封处理后, 荒漠草原杂草类占比 59.66%, 蒿类占比降至 29.99%, 豆科占比 5.19%, 禾本科占比 3.43%, 占比最小的是莎草科(1.73%)。围封并未显著地改变高山草原的功能群组成, 其中杂草类在围封内、外的占比分别为 65.31%和 68.52%, 然后为禾本科(16.38% 和15.01%)、莎草科(13.49%和 11.54%)和蒿类(2.71%和2.52%), 最后为豆科, 占比分别为 2.10%和 2.40%。
6 年围封显著增加了荒漠草原杂草类植物的覆盖度(表 2), 同时降低了蒿类和禾本科植物的覆盖度。2017 和 2018 年的围封显著地改变蒿类的覆盖度, 禾本科仅在 2018 年覆盖度呈现显著的变化。围封对高山草原功能群组成的影响没有显著的年际间差异。
2.2 围封对群落稳定性的影响
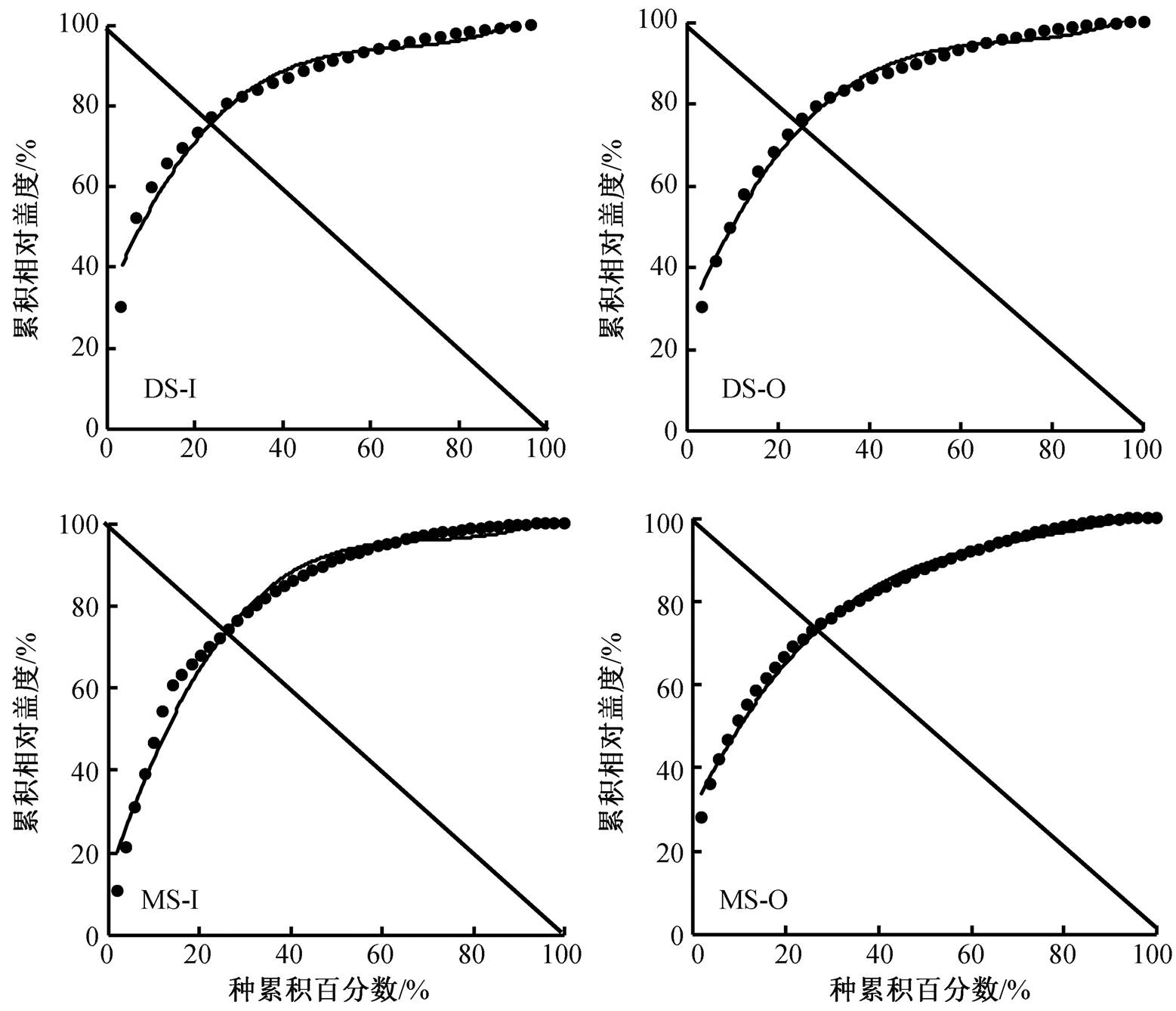
根据 Gordon 稳定性计算方法, 分别对高山草原、荒漠草原围栏内外植被进行平滑曲线拟合(图2), 曲线方程与 y=100−x 的交点坐标(表 3)表明, 对于不同植被类型的生态系统, 围封均增加其稳定性: 围封 6 年后荒漠草原围栏内交点坐标为(23.95, 76.05), 对应欧氏距离为 5.59, 小于围栏外欧氏距离7.21; 高山草原围栏内交点坐标为(26.26, 73.74), 欧氏距离为8.86, 亦小于放牧区欧氏距离10.18。
在荒漠草原围栏内, 2013—2018 年与稳定点(20, 80)的欧氏距离分别为 6.35, 1.90, 3.85, 2.91, 5.79 和 5.59, 稳定性呈现年际波动的趋势; 围栏外为 11.56, 6.36, 7.82, 7.35, 7.04 和 7.21, 除 2013 年外, 稳定性逐年增加并趋于稳定, 两者差值分别为 5.21, 4.46, 3.97, 4.44, 1.25 和 1.62。围封植被的稳定性均大于放牧处理, 且随围封年限增加, 围栏内外的稳定性差距有缩小趋势; 对于高山草原, 围栏内 2013 —2018 年欧氏距离分别为 10.62, 8.15, 7.82, 7.35, 7.04 和 7.21, 稳定性减小后趋于稳定, 放牧处理下欧氏距离为 11.21, 10.44, 9.47, 9.76, 8.61 和 10.18, 两者差值为 0.58, 2.29, 0.01, −0.22, 3.60 和 1.32。围封内外的稳定性差异无明显的年际间变化规律。
2.3 群落组成与Gordon稳定性的关系
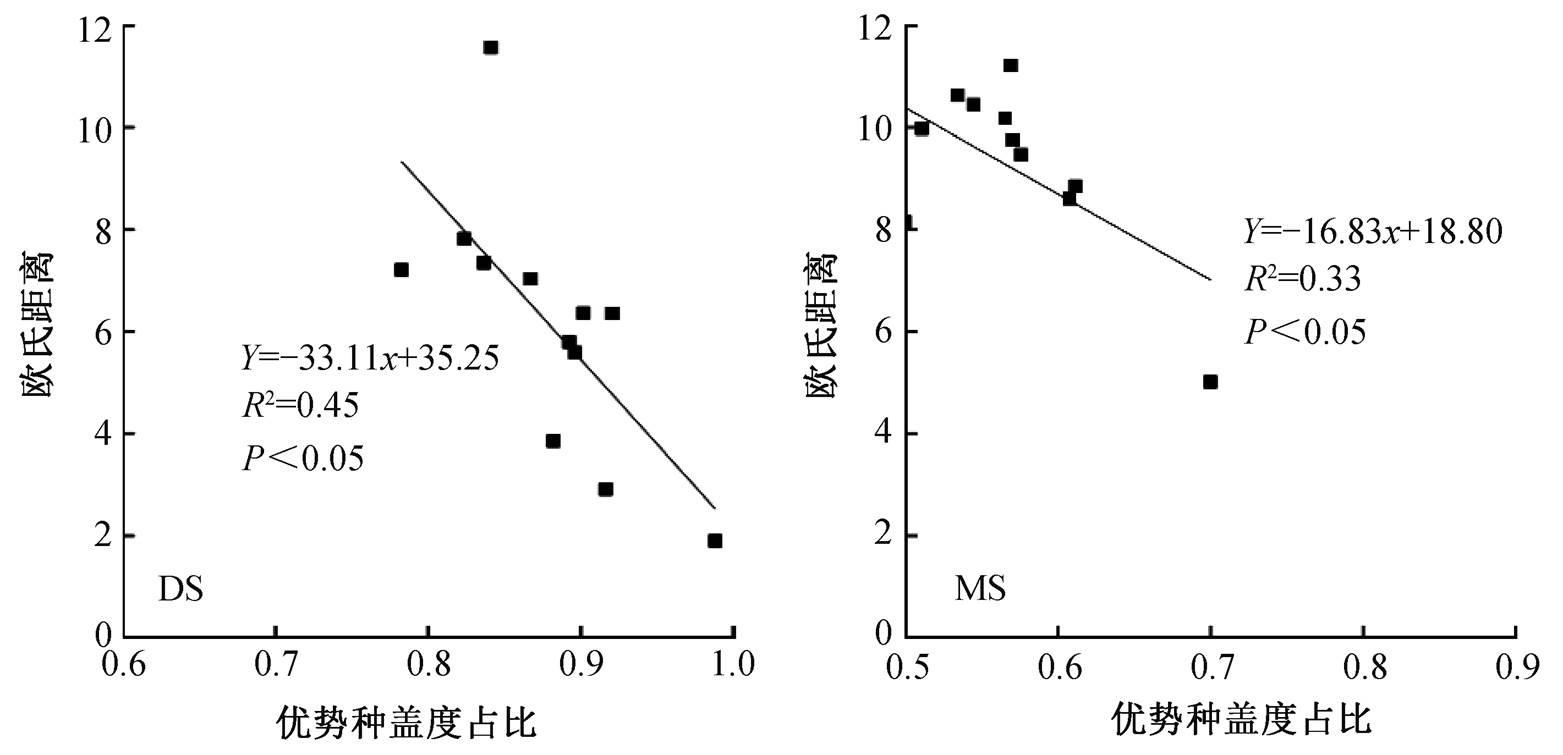
将 Gordon 稳定性中的欧氏距离与优势种盖度占比(反映群落组成状况)间建立相关关系(图 3), 我们发现无论是荒漠草原还是高山草原, 欧式距离和优势种盖度占比均呈显著的负相关关系, 即优势种盖度占群落的比例越高, 欧式距离越小, 即群落稳定性越高。同时发现, 荒漠草原优势种盖度占比在围栏内外及各年均高于高山草原对应情景的优势种盖度占比。
表2 2013—2018年围封对不同植物功能群盖度的影响
Table 2 Enclosure effects on the cover of different plant functional groups from 2013 to 2018
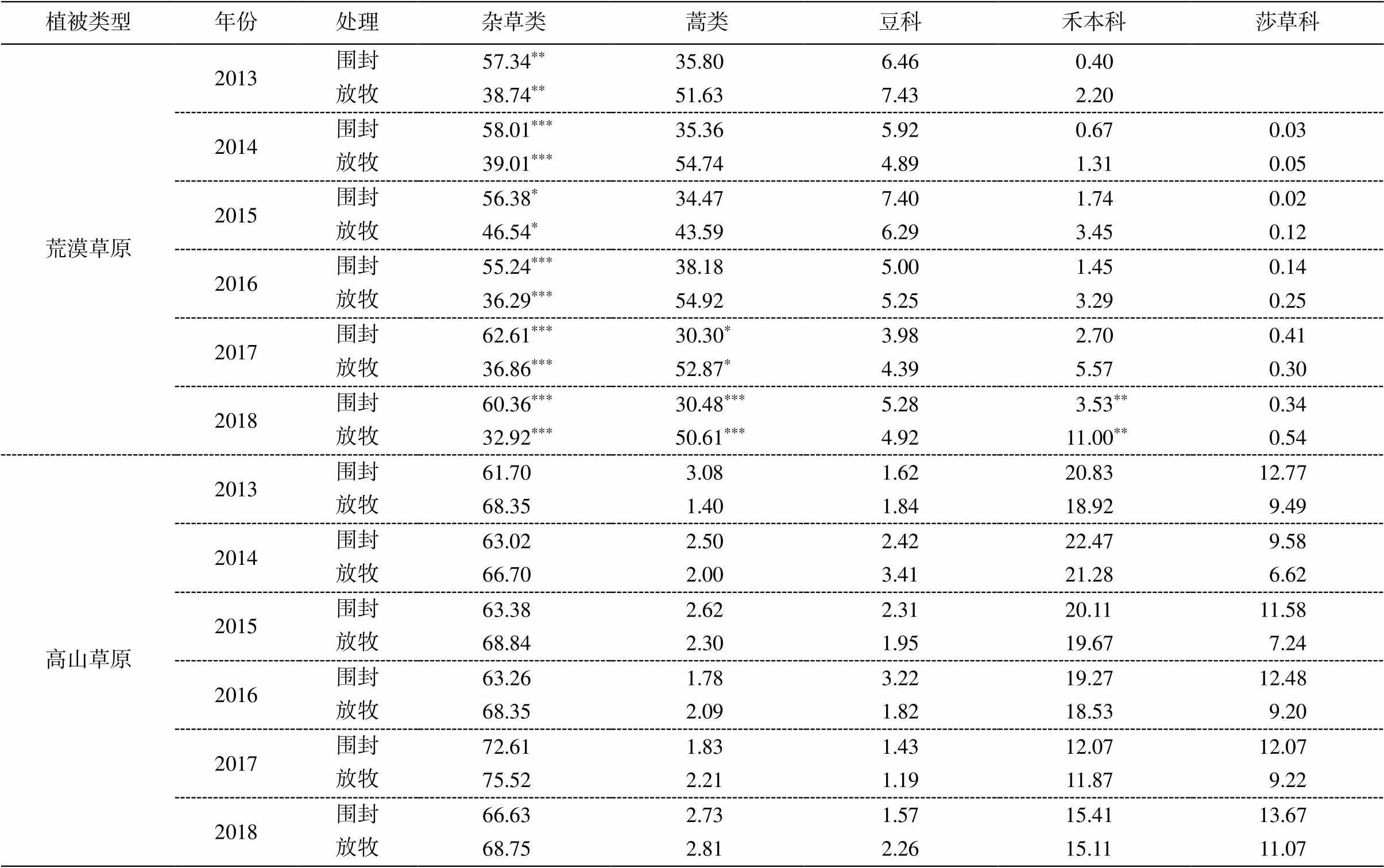
植被类型年份处理杂草类蒿类豆科禾本科莎草科 荒漠草原2013围封57.34**35.806.460.40 放牧38.74**51.637.432.20 2014围封58.01***35.365.920.670.03 放牧39.01***54.744.891.310.05 2015围封56.38*34.477.401.740.02 放牧46.54*43.596.293.450.12 2016围封55.24***38.185.001.450.14 放牧36.29***54.925.253.290.25 2017围封62.61***30.30*3.982.700.41 放牧36.86***52.87*4.395.570.30 2018围封60.36***30.48***5.283.53**0.34 放牧32.92***50.61***4.9211.00**0.54 高山草原2013围封61.703.081.6220.8312.77 放牧68.351.401.8418.929.49 2014围封63.022.502.4222.479.58 放牧66.702.003.4121.286.62 2015围封63.382.622.3120.1111.58 放牧68.842.301.9519.677.24 2016围封63.261.783.2219.2712.48 放牧68.352.091.8218.539.20 2017围封72.611.831.4312.0712.07 放牧75.522.211.1911.879.22 2018围封66.632.731.5715.4113.67 放牧68.752.812.2615.1111.07
表3 2013—2018年高山草原和荒漠草原围栏内外的稳定性
Table 3 Community stability of mountain steppe and dry steppe inside and outside enclosure from 2013 to 2018
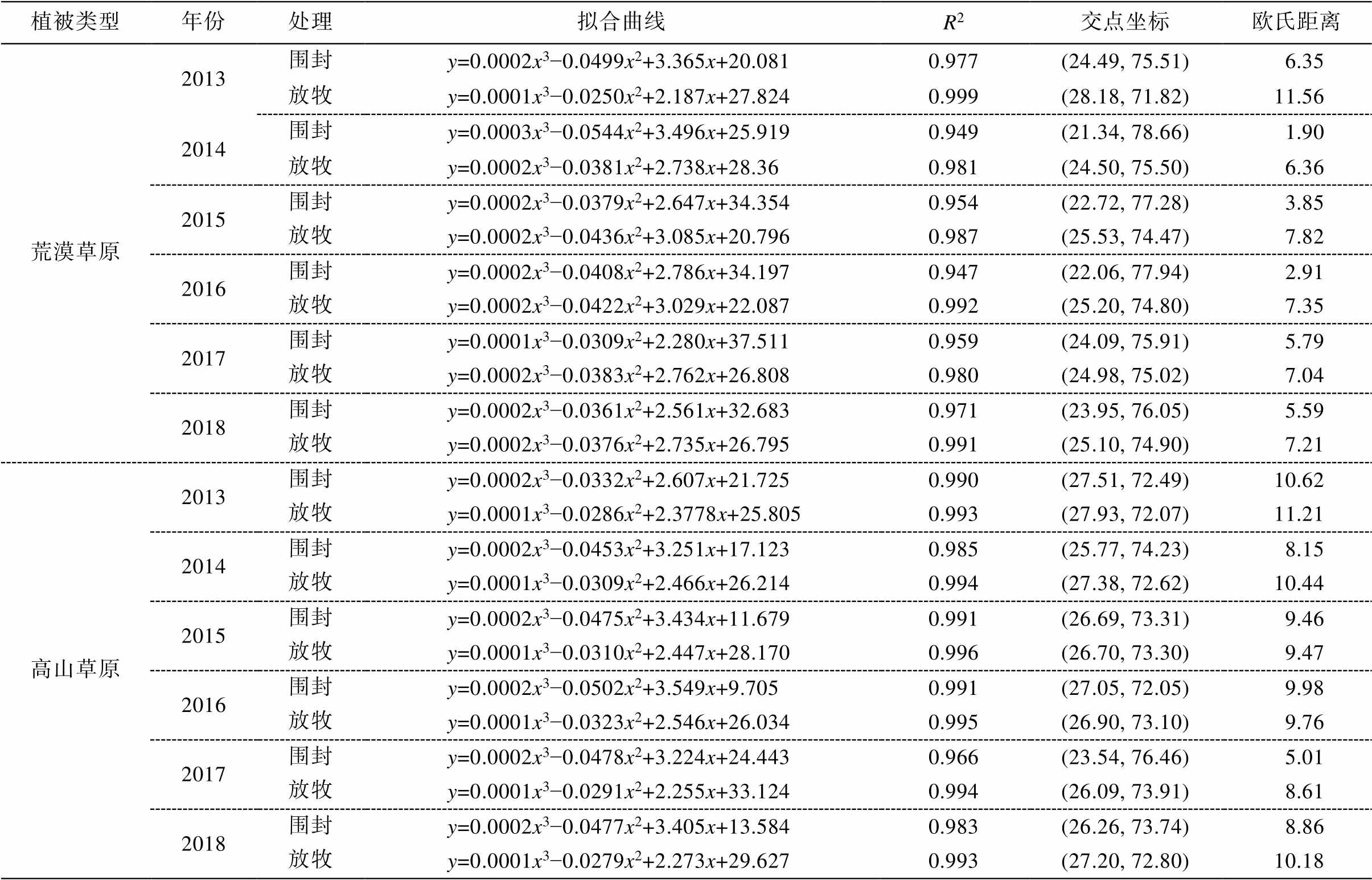
植被类型年份处理拟合曲线R2交点坐标欧氏距离 荒漠草原2013围封y=0.0002x3−0.0499x2+3.365x+20.0810.977(24.49, 75.51)6.35 放牧y=0.0001x3−0.0250x2+2.187x+27.8240.999(28.18, 71.82)11.56 2014围封y=0.0003x3−0.0544x2+3.496x+25.9190.949(21.34, 78.66)1.90 放牧y=0.0002x3−0.0381x2+2.738x+28.360.981(24.50, 75.50)6.36 2015围封y=0.0002x3−0.0379x2+2.647x+34.3540.954(22.72, 77.28)3.85 放牧y=0.0002x3−0.0436x2+3.085x+20.7960.987(25.53, 74.47)7.82 2016围封y=0.0002x3−0.0408x2+2.786x+34.1970.947(22.06, 77.94)2.91 放牧y=0.0002x3−0.0422x2+3.029x+22.0870.992(25.20, 74.80)7.35 2017围封y=0.0001x3−0.0309x2+2.280x+37.5110.959(24.09, 75.91)5.79 放牧y=0.0002x3−0.0383x2+2.762x+26.8080.980(24.98, 75.02)7.04 2018围封y=0.0002x3−0.0361x2+2.561x+32.6830.971(23.95, 76.05)5.59 放牧y=0.0002x3−0.0376x2+2.735x+26.7950.991(25.10, 74.90)7.21 高山草原2013围封y=0.0002x3−0.0332x2+2.607x+21.7250.990(27.51, 72.49)10.62 放牧y=0.0001x3−0.0286x2+2.3778x+25.8050.993(27.93, 72.07)11.21 2014围封y=0.0002x3−0.0453x2+3.251x+17.1230.985(25.77, 74.23)8.15 放牧y=0.0001x3−0.0309x2+2.466x+26.2140.994(27.38, 72.62)10.44 2015围封y=0.0002x3−0.0475x2+3.434x+11.6790.991(26.69, 73.31)9.46 放牧y=0.0001x3−0.0310x2+2.447x+28.1700.996(26.70, 73.30)9.47 2016围封y=0.0002x3−0.0502x2+3.549x+9.7050.991(27.05, 72.05)9.98 放牧y=0.0001x3−0.0323x2+2.546x+26.0340.995(26.90, 73.10)9.76 2017围封y=0.0002x3−0.0478x2+3.224x+24.4430.966(23.54, 76.46)5.01 放牧y=0.0001x3−0.0291x2+2.255x+33.1240.994(26.09, 73.91)8.61 2018围封y=0.0002x3−0.0477x2+3.405x+13.5840.983(26.26, 73.74)8.86 放牧y=0.0001x3−0.0279x2+2.273x+29.6270.993(27.20, 72.80)10.18
3 讨论
我们发现, 在荒漠草原, 围封降低了喜食性植物功能群(如禾草和莎草)在群落中的比重。这与一般的认识是相反的, 即食草者会优先选择优良的牧草[26]。主要原因是, 在荒漠草原, 杂草是群落中的优势功能群, 禾草和莎草在群落中的占比很低。此时动物为了生存优先采食临近的杂草, 而不大可能消耗过多的体力寻找优质的牧草。当围栏外的杂草植物覆盖度减少时, 次优势种蒿类占据空余的生存空间。研究发现, 一些蒿草物种具有强大的繁殖能力, 并且会抑制周围优良牧草的生长[16]。在高山草原, 围封对群落组成的影响较弱。相对于荒漠草原, 高山草原拥有更充沛的降水, 可以在一定程度上调控放牧对群落组成的影响[27–28]。在不同降水条件下, 不同植被类型对围封的响应是有差异的[29]。在荒漠草原, 短期放牧对群落组成的影响在很大程度上是随机的。随着围封年限的增加, 放牧对群落中优势种盖度的影响逐渐增强。围栏外群落发生着由优势种向次优势种演替的进程。这说明短期放牧和长期放牧对群落组成的影响有差异[29–30]。然而, 在高山草原, 围封时间没有明显地影响群落组成对围封的响应。
在荒漠草原和高山草原, 围封增加了群落的Gordon 稳定性。Gordon 稳定性主要由群落中优势种的占比决定。我们建立了优势种占比与稳定性的定量关系, 结果表明 Gordon 稳定性欧氏距离与群落中优势种盖度占比呈显著的负相关关系。当优势种在群落中占比较大时, 群落结构相对稳定。对于荒漠草原, 由于杂草类植物是优势功能群, 在围栏内的相对盖度远远大于围栏外, 导致围栏内群落稳定性更高, 同时蒿草类功能群在围栏外的相对盖度更大。研究发现, 对于高寒地区的草地, 以蒿类为主的植物群落往往稳定性更低[31], 即蒿类植物盖度过多不利于群落稳定。可能的原因是蒿类植物的分布较为集中, 更耐踩踏, 易适应围栏外的放牧环境[16]。对于高山草原, 优势种杂草类植物在围栏内外差异不大, 在牧草资源丰富的情况下, 植食者会优先选择优质的牧草。围封有助于喜食性牧草的生长, 进而增强群落的稳定性。对于荒漠草原, 围封年限的增加会减小围栏内外稳定性的差异, 说明围栏外的物种可能不断向着优势种的方向进化。对于高山草原, 围栏内外稳定性的差异受围封年限的影响较弱。草原群落的稳定性不仅与群落组成紧密相关, 还可能受土壤养分、气候条件、外界干扰等因素调控[18,32]。此外, 我们发现荒漠草原群落的Gordon 稳定性高于高山草原, 其直接原因为荒漠草原优势种在群落中的占比高于高山草原。高山草原虽然植物覆盖度高, 但各植物功能群分布较平均。
4 结论
本文结果表明, 围封对于不同草地类型群落组成和稳定性影响迥异, 在荒漠草原, 围封显著地提高其稳定性, 但对于水土条件较好的高山草原, 围封并未显著地改善其群落稳定性。尽管围封作为促进草原恢复的主要措施, 具有高效、简便和易推广的优势, 但对于不同类型的草地生态系统, 应因地制宜地开展恢复措施。围封前应做充足的调查研究, 从土壤结构、气候条件、植被类型和放牧强度等多个维度考虑围封的可行性及其潜在影响, 不能一概而论。同时, 辅以适当休牧、轮牧或季节性放牧等措施, 才能真正实现草原生态系统服务的可持续利用, 降低管理和恢复成本, 将草场的生态学效益和经济学效益发挥到最大程度。
参考文献
[1] Suttie J M, Reynolds S G, Batello C, et al. Grasslands of the world. Rome: FAO, 2005
[2] 董全民, 赵新全, 马玉寿, 等. 牦牛放牧强度对小嵩草草甸两季轮牧草场植物群落数量特征的影响. 生态学杂志, 2011, 30(10): 2233–2239
[3] Briske D D, Zhao M, Han G, et al. Strategies to alleviate poverty and grassland degradation in Inner Mongolia: intensification vs production efficiency of livestock systems. Journal of Environmental Manage-ment, 2015, 152: 177–182
[4] 李西良, 侯向阳, 吴新宏, 等. 草甸草原羊草茎叶功能性状对长期过度放牧的可塑性响应. 植物生态学报, 2014, 38(5): 440–451
[5] Zhang M, Zhang L, Zhang Y, et al. Pastureland transfer as a livelihood adaptation strategy for herds-men: a case study of Xilingol, Inner Mongolia. The Rangeland Journal, 2017, 39(2): 179–187
[6] Niu K, Messier J, He J S, et al. The effects of grazing on foliar trait diversity and niche differentiation in Tibetan alpine meadows. Ecosphere, 2015, 6(9): 1–15
[7] 苏军虎, 南志标, 纪维红. 家畜放牧对草地啮齿动物影响的研究进展. 草业学报, 2016, 25(11): 136–148
[8] Zheng S X, Ren H Y, Lan Z C, et al. Effects of grazing on leaf traits and ecosystem functioning in Inner Mongolia grasslands: scaling from species to community. Biogeosciences, 2010, 7(3): 1117–1132
[9] Jing Z B, Cheng J M, Su J S, et al. Changes in plant community composition and soil properties under 3-decade grazing exclusion in semiarid grassland. Eco-logical Engineering, 2014, 64: 171–178
[10] 德米特里耶夫A M. 草地经营附草地学基础. 蔡元定等, 译. 北京: 财政经济出版社, 1955
[11] 刘菊红, 王忠武, 郝敦元, 等. 重牧对荒漠草原主要植物种和功能群组织力的影响. 中国草地学报, 2018, 40(5): 85–92
[12] 张婧, 左小安, 吕朋, 等. 科尔沁沙地草地植被对围封和放牧的响应. 中国沙漠, 2017, 37(3): 446–452
[13] 薛亚芳, 宗宁, 何念鹏, 等. 长期围封和自由放牧对高寒草甸土壤微生物群落结构及碳源代谢多样性的影响. 应用生态学报, 2018, 29(8): 2705–2712
[14] 李璠, 周国英, 杨路存, 等. 围栏封育对青海湖流域主要植物群落多样性与稳定性的影响. 水土保持研究, 2013, 20(4): 135–140
[15] Weigelt A, Schumacher J, Roscher C, et al. Does biodiversity increase spatial stability in plant commu-nity biomass. Ecology Letters, 2008, 11: 338–347
[16] 罗久富, 周金星, 赵文霞, 等. 围栏措施对青藏高原高寒草甸群落结构和稳定性的影响. 草业科学, 2017, 34(3): 565–574
[17] Pimm S L. The complexity and stability of ecosys-tems. Nature, 1984, 307: 321–326
[18] Tilman D, Reich P B, Knops J M H. Biodiversity and ecosystem stability in a decade-long grassland experi-ment. Nature, 2006, 441: 629–632
[19] Poissonet P S, Poissonet J A, Gordon M P, et al. A comparison of sampling methods in dense herbacea-ous pasture. Rangeland Ecology & Management — Journal of Range Management Archives, 1973, 26(1): 65–67
[20] Grime J P. Benefits of plant diversity to ecosystems: immediate, filter and founder effects. Journal of Ecology, 1998, 86(6): 902–910
[21] Li S G, Asanuma J, Eugster W, et al. Net ecosystem carbon dioxide exchange over grazed steppe in central Mongolia. Global Change Biology, 2005, 11(11): 1941–1955
[22] Fernandezgimenez M, Allendiaz B. Vegetation change along gradients from water sources in three grazed Mongolian ecosystems. Plant Ecology, 2001, 157(1): 101–118
[23] 郑元润. 森林群落稳定性研究方法初探. 林业科学, 2000, 36(5): 28–32
[24] 张瑜, 王普昶, 莫本田, 等. 不同放牧强度下贵州喀斯特草地植被空间特征与生产力相关性分析. 草业与畜牧, 2015(4): 31–36
[25] 李育中. 植物群落稳定性的一种测定方法. 中国草地, 1991(2): 78–80
[26] Diaz S, Lavorel S, McIntyre S U E, et al. Plant trait responses to grazing — a global synthesis. Global Change Biology, 2007, 13(2): 313–341
[27] Guo T, Lohmann D, Ratzmann G, et al. Response of semi-arid savanna vegetation composition towards grazing along a precipitation gradient — the effect of including plant heterogeneity into an ecohydrological savanna model. Ecological Modelling, 2016, 325: 47–56
[28] Koerner S E, Collins S L, Blair J M, et al. Rainfall variability has minimal effects on grassland recovery from repeated grazing. Journal of Vegetation Science, 2014, 25(1): 36–44
[29] Xiong D, Shi P, Zhang X, et al. Effects of grazing exclusion on carbon sequestration and plant diversity in grasslands of China — a meta-analysis. Ecological Engineering, 2016, 94: 647–655
[30] 刘玉, 刘振恒, 邓蕾, 等. 季节性放牧对草地植物多样性与功能群特征的影响. 草业科学, 2016, 33(7): 1403–1409
[31] 杨力军, 李希来, 石德军, 等. 青藏高原“黑土滩”退化草地植被演替规律的研究. 青海草业, 2005(1): 2–5
[32] Wen D, He N, Zhang J. Dynamics of soil organic carbon and aggregate stability with grazing exclusion in the Inner Mongolian grasslands. Plos One, 2016, 11(1): e0146757
Effects of Grazing Exclusion on the Vegetation Community Composition and the Community Stability of Dry Steppe and Mountain Steppe Ecosystems in Mongolia
LIAO Hanru, Indree Tuvshintogtokh, GUO Tong†, ZHAO Jingxue
College of Urban and Environmental Sciences, Laboratory for Earth Surface Processes of the Ministry of Education, Peking University, Beijing 100871; † Corresponding author, E-mail: tongg@pku.edu.cn
Abstract Two types of typical steppe ecosystems in Mongolia (dry steppe and mountain steppe) with two treatments (enclosure and grazing) were studied to analyze the vegetation cover from 2013 to 2018. The plant species of the community were divided into five functional groups: forbs, artemisia, legumes, grasses and sedges. The Gordon stability index was used to evaluate the effect of enclosure on the community stability. Here are the results. 1) Enclosure significantly increased the coverage of forbs in the dry steppe, while reduced the coverage of artemisia and gramineous groups. The effects had no obvious variations among years. For the mountain steppe, enclosure slightly affected the coverage of different functional groups. 2) Enclosure increased the stability of the dry steppe while little influenced the stability of the mountain steppe. 3) The Gordon stability index had a significantly positive relation with the cover ratio of dominant species to the communities. The ratio in the dry steppe was higher than that in the mountain steppe, leading to a higher community stability of the dry steppe. Before the enclosure management in grasslands, we should fully consider local environmental conditions and the potential impact of enclosure on the plant community and the ecosystem. At the same time, grazing rest, rotational grazing, seasonal grazing and other measures should be supplemented to improve the use value of grassland ecosystem services.
Key words inter-annual variability; community structure; functional groups; Gordon stability; vegetation cover
doi: 10.13209/j.0479-8023.2020.017
国家自然科学基金(31621091)资助
收稿日期: 2019-03-22;
修回日期: 2019-04-23
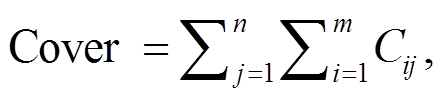 (1)
(1)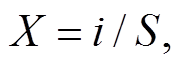 (2)
(2)