我国东部典型森林木本植物的气孔特征及其对氮添加的响应
姜星星 邹安龙 王媛媛 周序力 吉成均 †
北京大学城市与环境学院生态学系, 北京 100871
摘要 利用我国东部森林养分添加实验平台, 测定18种优势木本植物叶片的气孔特征。结果表明, 不同生活型木本植物的气孔特征有显著的差异( P <0.05), 我国东部优势木本植物的气孔特征具有明显的纬度格局, MAT, MAP和PET是重要的影响因子。各优势树种气孔特征之间存在显著的相关性, 气孔长度和气孔密度显著负相关, 气孔导度则随着气孔长度的增加而增加, 随着气孔密度的增加而降低。氮添加对不同物种气孔特征的影响不同, 不同温度带和生活型木本植物的气孔特征对氮添加的响应也存在差异。
关键词 氮添加; 气孔特征; 森林; 叶片
气孔是植物与环境进行水分和气体交换的主要通道, 直接影响植物的蒸腾作用和光合作用 [1] 。气孔不但会影响植物水分利用和营养吸收等过程, 对整个生态系统也有重要的调节作用 [2–3] 。为适应复杂多变的环境, 植物一方面通过气孔的打开和闭合, 做出快速反应, 调整气孔导度来应对短期的环境变化; 另一方面则在长期与环境胁迫抗争的过程中, 通过改变气孔的大小和密度来适应环境变化 [4] 。因此, 研究植物气孔是理解植物对环境变化适应机制的有效手段之一。目前, 针对气孔的研究主要集中于气孔对水分 [5] 、光质和光强 [6] 、温度 [7] 及CO 2 浓度 [8–9] 等因素的响应方面, 而对氮添加响应的研究较少。
氮沉降已成为重大的环境问题之一, 影响着森林生态系统的养分可利用状况和植物的生长 [10–11] 。氮素是影响植物气孔特征的重要营养元素, 一方面, 氮素是植物体内1,5-二磷酸核酮糖羧化酶/加氧酶(Rubisco)和叶绿素的重要组成成分, 可以通过调控叶绿素含量、酶含量及酶活性等影响光合作用, 进而影响植物叶片的气孔特征 [12] 。另一方面, 氮素与脱落酸(ABA)的积累有密切的联系, 而ABA是介导根冠间信息传递的核心信号物质, 因此氮素可通过调控根冠间信息传递而影响气孔导度 [13] 。已有研究发现, 氮添加会显著影响植物叶片的某些功能属性, 如施肥可增加叶片氮含量, 改变元素的比例关系, 提高叶绿素含量和净光合速率, 增强植物光合能力等 [14–17] 。虽然气孔在叶片功能中具有重要作用, 但在解释这些功能变化时, 往往缺少气孔特征方面的数据支持。因此, 分析气孔特征在不同养分条件下的变化规律, 可为探索植物对氮沉降的响应机制提供新的有力证据。
研究区域尺度植物功能属性及其与环境变化的关系, 对于理解植物对环境的适应机制及生态模型的构建有重要意义 [18–21] 。在区域尺度探讨森林对氮沉降的响应已成为研究热点, 目前主要集中在植被生物量、生物多样性、资源分配等方面 [16,22–23] ,缺少对大尺度格局上植物气孔特征对氮添加响应规律的探讨。本实验利用中国森林养分添加实验平台(Nutrient Enrichment Experiments in Chinese Forests, NEECF) [23] , 沿纬度梯度(18°—51°N)在中国东部森林7个地点设置9个施肥样地, 分析优势树种的气孔特征, 探讨以下科学问题: 1)优势树种的气孔特征及其纬度格局; 2)优势树种气孔特征对氮添加的响应规律。
1 材料与方法
1.1 研究平台
NEECF平台利用长期野外定位模拟氮沉降法, 研究氮沉降对中国东部典型森林生态系统的影响。平台包含中国东部的主要森林生态系统, 从北到南分别为内蒙古根河(GH)寒温带落叶松原始林、黑龙江五营(WY)温带阔叶红松原始林、河北塞罕坝(SHB)落叶松人工林、北京东灵山(DLS)温带辽东栎和白桦次生林、安徽牯牛降(GNJ)亚热带常绿阔叶原始林、福建武夷山(WYS)亚热带常绿阔叶原始林、海南尖峰岭(JFL)热带山地雨林原始林和次生林。本研究涉及9个样地, 每个样地设置9或12个样方(3~4个施肥梯度, 3个重复样方, 样方大小为20 m×20 m)。各样地的基本信息见表1。
1.2 样品采集
样品采集时间为2016年5—8月, 共采集9个样地的优势或常见木本植物18种(表1)。对每种植物, 在每一样方内选取3株年龄相近的成熟健康植株, 利用人工爬树或高枝剪的方法, 采集树冠中上部发育程度相近、位置相似的健康带叶片枝条, 采下后立即用LI-6400便携式光合仪测量叶片的气孔导度(stomatal conductance), 然后放入冰盒保存。带回实验室后, 在每个枝条上选取3个形态相近的健康成熟叶片, 放入FAA固定液中固定, 并置于4ºC冰箱中保存。气孔导度表示气孔张开的程度, 是反应植物与大气间气体通量的重要参数, 影响植物的光合作用和蒸腾作用 [24] 。
表1 NEECF实验平台基本情况
Table 1 General information of NEECF sites
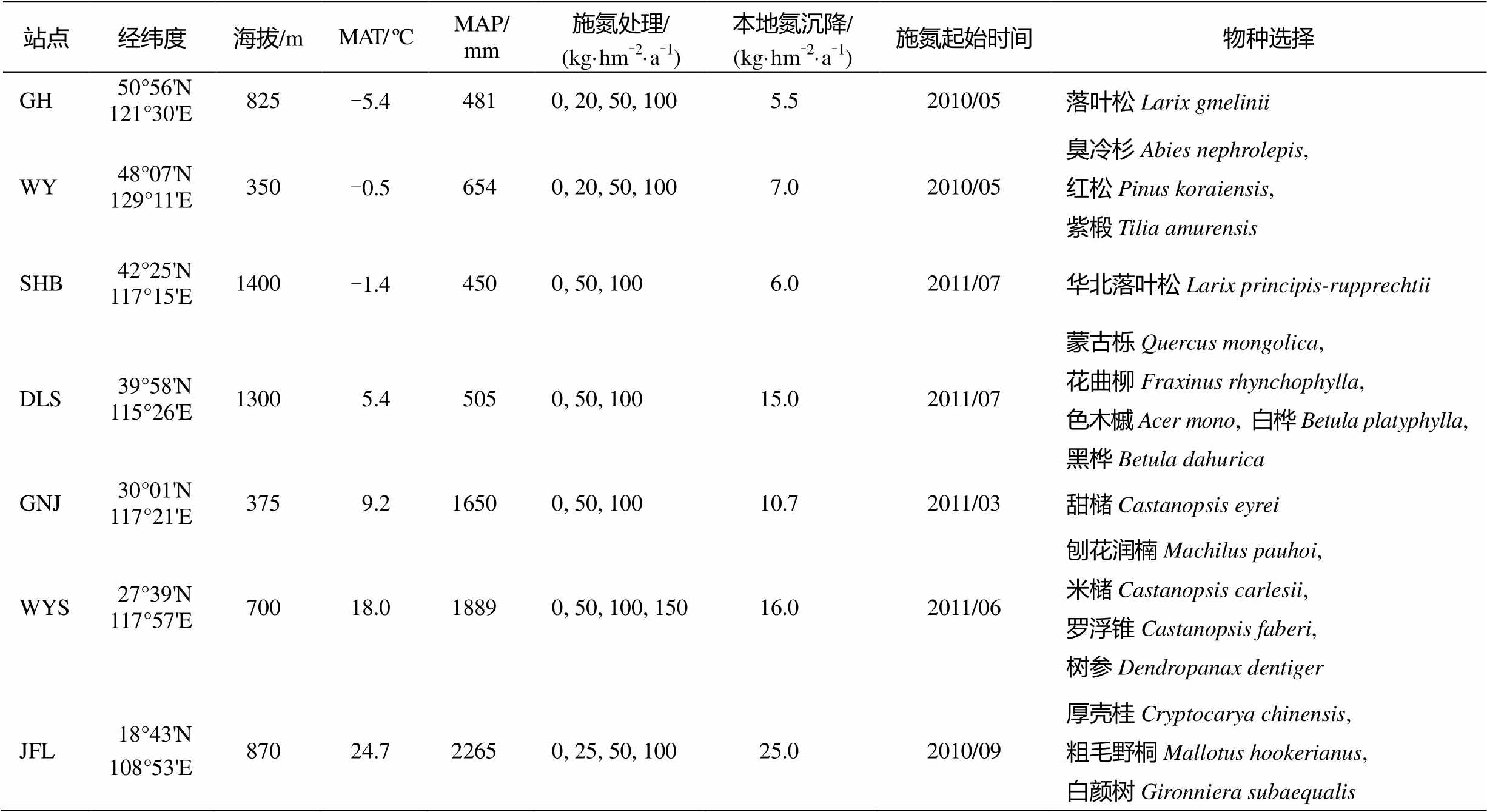
站点经纬度海拔/mMAT/ ºCMAP/ mm施氮处理/ (kg·hm-2·a-1)本地氮沉降/ (kg·hm-2·a-1)施氮起始时间物种选择 GH50°56'N121°30'E825-5.44810, 20, 50, 1005.52010/05落叶松Larix gmelinii WY48°07'N129°11'E350-0.56540, 20, 50, 1007.02010/05臭冷杉Abies nephrolepis, 红松Pinus koraiensis, 紫椴Tilia amurensis SHB42°25'N117°15'E1400-1.44500, 50, 1006.02011/07华北落叶松Larix principis-rupprechtii DLS39°58'N115°26'E13005.45050, 50, 10015.02011/07蒙古栎Quercus mongolica, 花曲柳Fraxinus rhynchophylla, 色木槭Acer mono, 白桦Betula platyphylla, 黑桦Betula dahurica GNJ30°01'N117°21'E3759.216500, 50, 10010.72011/03甜槠Castanopsis eyrei WYS27°39'N117°57'E70018.018890, 50, 100, 15016.02011/06刨花润楠Machilus pauhoi, 米槠Castanopsis carlesii, 罗浮锥Castanopsis faberi, 树参Dendropanax dentiger JFL18°43'N108°53'E87024.722650, 25, 50, 10025.02010/09厚壳桂Cryptocarya chinensis, 粗毛野桐Mallotus hookerianus, 白颜树Gironniera subaequalis
1.3 指标测量
采用指甲油印迹法, 获取叶片下表面气孔结构特征 [25] 。在距叶尖约2/3处的叶片下表面均匀涂抹一层指甲油, 在室温下自然晾干, 用透明胶带粘取叶片下表皮印记后粘在载玻片上, 制成临时装片, 利用MoticB5光学显微镜观察, Motic Images Plus 2.0显微成像系统照相和测定。
测定指标包括气孔长度(stomatal length)和气孔密度(stomatal density)。本研究以气孔器保卫细胞长轴长度代表气孔长度(μm), 用单位面积的气孔数除以对应的视野面积来计算气孔密度(个/mm 2 )。每片叶子选取4个视野进行气孔长度和气孔密度的计数和测量(避开叶脉区域), 每个视野随机选择5个气孔测量长度, 各指标均取4个视野的平均值。SPI指数(stomatal pore area index)是反映植物潜在光合能力的重要指标, 计算公式为: SPI=气孔长度 2 ×气孔密度×10 -4 , 可近似理解为单位面积的气孔面积 [26] 。
1.4 数据分析
采用简单线性回归分析纬度、气候因子和施氮处理对植物气孔特征的影响, 并探讨气孔特征之间的相关关系。用 t 检验方法, 分析不同生活型植物气孔特征的差异( α =0.05)。用Pearson相关分析探讨氮添加对不同物种气孔特征的影响。统计分析使用SPSS16.0完成, 图表绘制使用Sigmaplot11.0软件完成。
2 结果
2.1 不同生活型木本植物的气孔特征
不同生活型木本植物的气孔特征有显著的差异( P <0.05)。与阔叶乔木相比, 针叶乔木的气孔长度更长, 气孔密度更低, SPI指数较高, 阔叶与针叶乔木的气孔导度差异不显著。落叶乔木的气孔长度和SPI 指数显著高于常绿乔木, 但气孔导度显著低于常绿乔木, 二者的气孔密度无显著差别(图1)。
2.2 纬度和气候因子对气孔特征的影响
我国东部优势木本植物的气孔特征具有明显的纬度格局, 各物种的气孔长度随着纬度的增加而显著加大, 而各物种的气孔密度与纬度存在显著的负相关关系(蒙古栎除外, 图 2)。年均温(MAT)、年 降水量(MAP)和潜在蒸散量(PET)是影响气孔特征的重要气候因子(图 3)。随着年均温的增加, 气孔长度显著降低, 而各物种的气孔密度显著增加(蒙古栎除外)。年降水量与各物种的气孔长度呈现显著的负相关关系, 而与气孔密度显著正相关(蒙古栎除外)。各物种的气孔长度随着潜在蒸散量的增加而显著降低, 气孔密度则随着潜在蒸散量的增加而显著增加(蒙古栎除外)。

图1 不同生活型植物气孔特征箱线图
Fig. 1 Box plots of stomatal traits across different lifeform
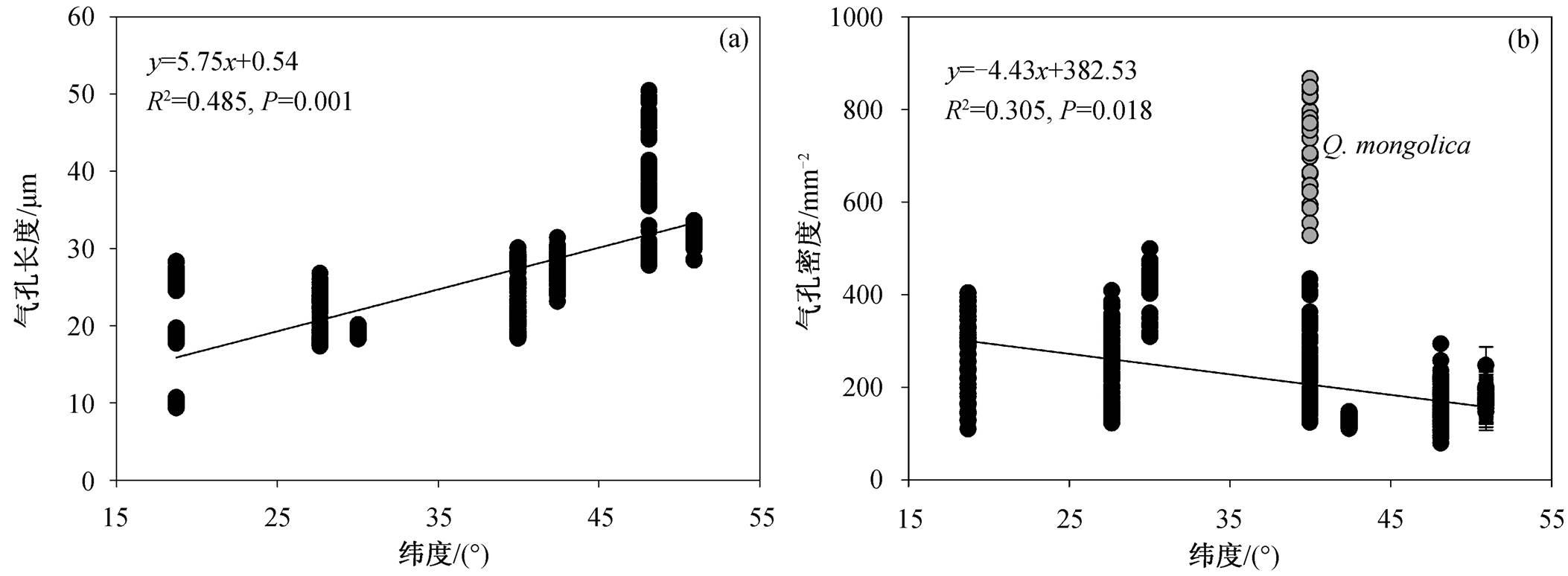
图2 气孔长度和气孔密度的纬度格局
Fig. 2 Latitudinal patterns of stomatal length and stomatal density
2.3 气孔特征之间的相关关系
如图4所示, 我国东部森林优势树种各气孔特征之间存在显著的相关性( P <0.05)。在气孔指标之间, 气孔长度与气孔密度显著负相关, 气孔导度随着气孔长度的增加而增加, 随着气孔密度的增加而降低。
2.4 施氮对气孔特征的影响
氮添加对不同物种气孔特征的影响不同(图5)。18种木本植物中, 4种植物(紫椴、中龄林华北落叶松、色木槭和厚壳桂)的气孔长度与施氮浓度显著正相关, 4种植物(蒙古栎、米槠、粗毛野桐和白颜树)的气孔长度与氮添加显著负相关, 其余植物的气孔长度与施氮浓度关系不显著。气孔密度随着施氮浓度增加而增加的植物有5种(紫椴、刨花润楠、米槠、罗浮锥和白颜树), 呈负相关关系的有4种(红松、黑桦、甜槠和厚壳桂)。随着施氮浓度的增加, 3种植物(花曲柳、色木槭和刨花润楠)的气孔导度和4种植物(紫椴、中龄林华北落叶松、刨花润楠和罗浮锥)的SPI指数显著增加, 呈负相关关系的物种数均有3种, 分别为紫椴、黑桦和树参, 以及红松、黑桦和甜槠。
不同温度带和生活型木本植物的气孔特征对氮添加的响应不同(表2)。亚热带地区优势乔木的气孔导度随着施氮浓度的增加而增加, 热带地区木本植物叶片的气孔密度和SPI指数与施氮处理显著正相关。对针叶乔木而言, 施氮会抑制气孔导度的增加; 阔叶乔木则相反, 施氮会显著促进气孔导度的增加。施氮未显著影响落叶乔木的气孔特征( P > 0.05), 但显著促进常绿乔木气孔长度和气孔导度的增加( P <0.05)。综合考虑氮添加对全部木本植物的影响, 发现氮添加显著抑制气孔长度的伸长和SPI指数的增加, 但促进气孔导度的增加。
我们发现, 气孔长度与气孔密度的负相关关系(取对数后)在不同施氮处理下均存在(图6), 但气孔长度与气孔密度负相关关系拟合线的斜率在不同氮添加处理下存在差异, 随着施氮浓度的增加, 斜率降低。
3 讨论
3.1 不同生活型植物的气孔特征
叶片气孔特征在不同生活型木本植物间存在较大差异, 反映出不同生活型植物对环境适应的策略不同 [27] 。本研究发现, 与阔叶乔木相比, 针叶乔木的气孔具有更长的气孔长度和较低的气孔密度, 与已有的研究结果 [28–29] 一致。原因可能是, 由于针叶乔木多生长于贫瘠或冬季较寒冷地区, 往往受到生理干旱的影响 [30] , 气孔的低密度有利于提升植物的水分利用效率 [31] 。落叶乔木叶片的气孔长度显著大于常绿乔木, 可能是因为大的气孔有利于增强光合能力, 进而增加生物量的积累, 落叶乔木叶片为适应较短的生长周期而选择这种策略 [32] 。此外, 与常绿乔木相比, 落叶乔木叶片的气孔导度明显偏小, 这也是适应北方干燥气候的有效手段。针叶乔木和落叶乔木的SPI指数分别高于阔叶乔木和常绿乔木, 而叶片的SPI指数较高, 往往意味着叶片在单位面积上的光合能力较强 [26] 。因此, 我们推测针叶乔木和落叶乔木的生长潜力可能较高, 这也是针叶乔木应对北方冬季漫长的低温气候期以及落叶乔木应对较短的叶片寿命的有效适应策略 [19] 。
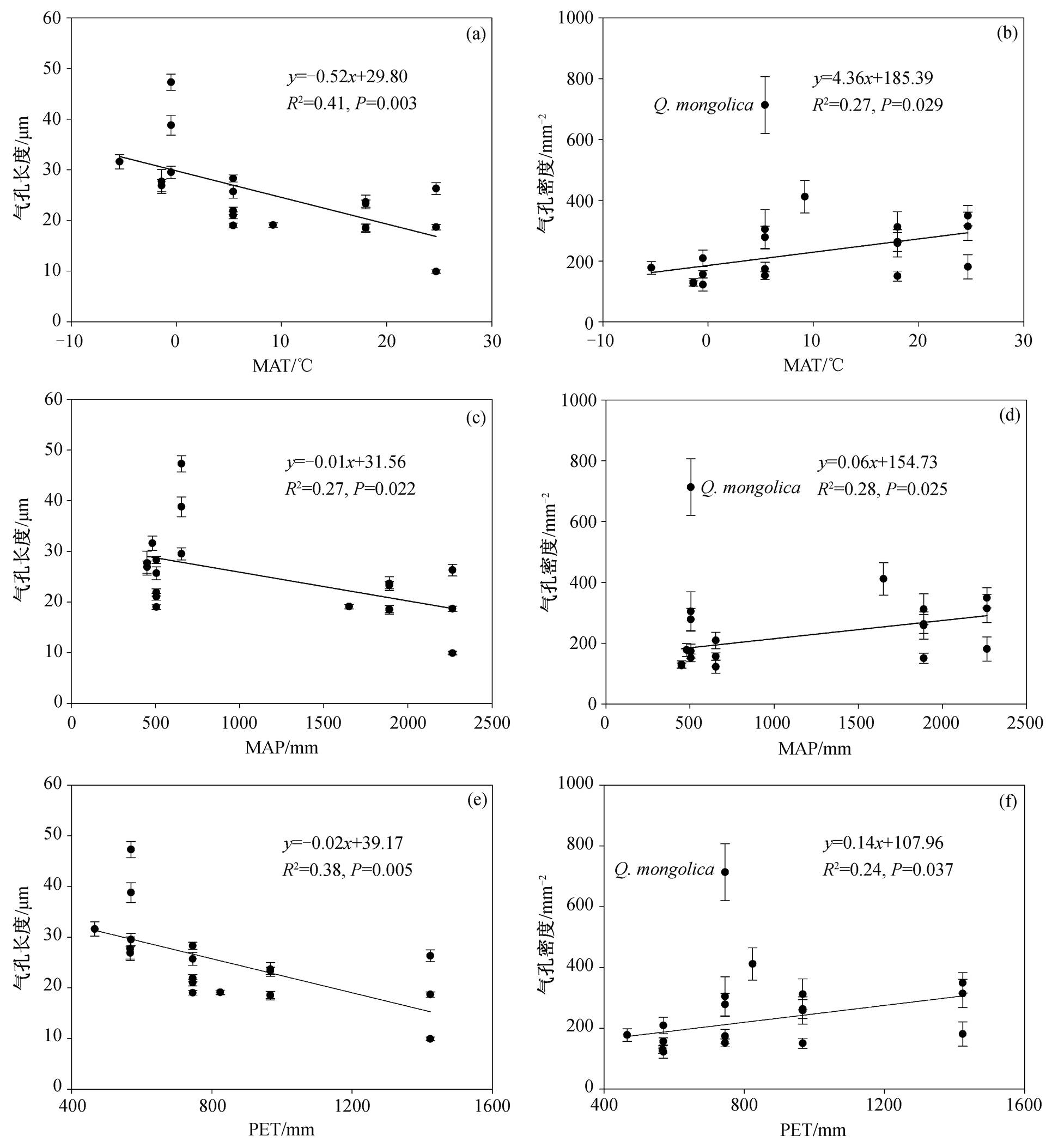
图3 气孔长度和气孔密度与气候因子的关系
Fig. 3 Changes in stomatal length and stomatal density with climate parameters
3.2 气孔特征的纬度格局及其对气候因子的响应
我国东部森林乔木树种叶片的气孔特征具有明显的纬度格局, 随着纬度的增加, 气孔长度显著增加, 气孔密度(蒙古栎除外)显著降低。为探讨产生这种格局的原因, 本研究选取气候因子MAT, MAP和PET分别分析其对气孔特征的影响, 其中, MAT和MAP数据引自文献[23], 该数据通过全国728个气象站点插值获取; PET引用WorldClim(http:// worldclim.org)的数据。研究发现, 气孔长度随着MAT的增加而显著降低, 气孔密度(蒙古栎除外)则相反。这可能是由于温度升高通常导致植物叶片的蒸腾作用加强, 植物会大量失水, 小而密的气孔有利于防止蒸腾作用导致的水分丧失 [33] 。气孔长度与MAP显著负相关, 与气孔密度(蒙古栎除外)则正相关, 可能是因为我国东部森林的MAT与MAP之间具有很强的自相关性, 这种格局反映气孔长度和气孔密度这两个指标最优化协调的结果 [4,34] 。此外, 随着PET的增加, 气孔向着小而密的方向发展, 小的气孔开、关迅速而灵活 [2] , 从而减少气孔打开的时间。因此, 这种策略是降低植物蒸腾作用的有效策略, 可减少水分的散失 [32] 。
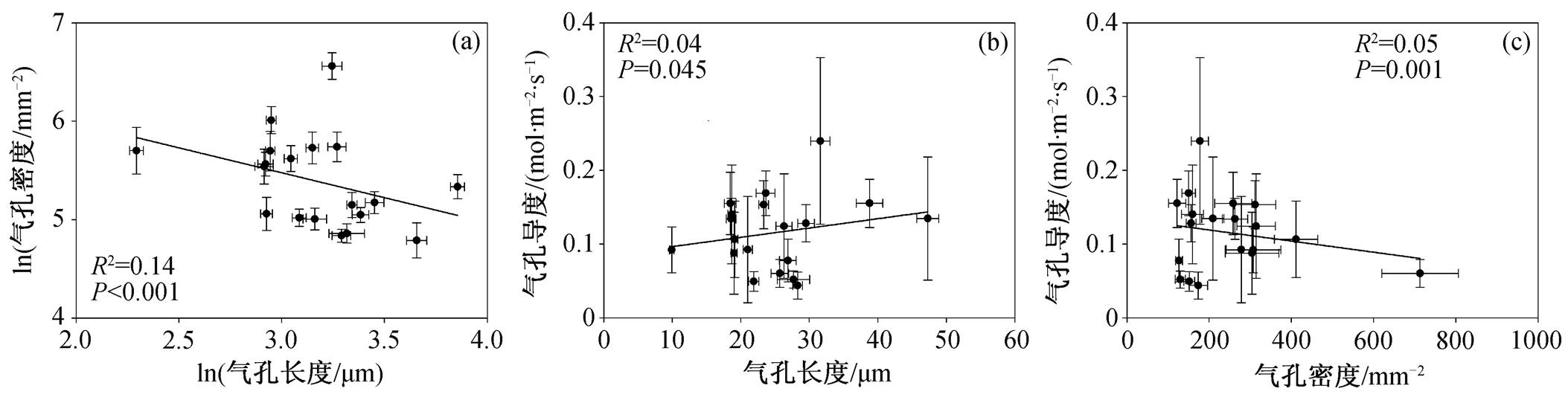
图4 气孔特征之间的相关关系
Fig. 4 Correlations among stomatal traits
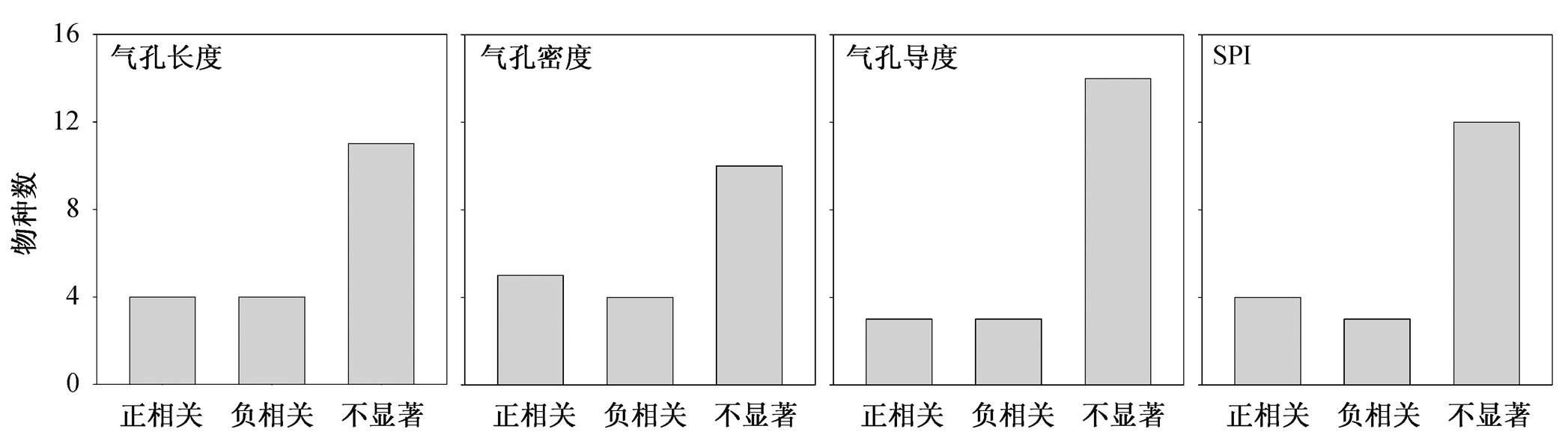
图5 施氮与不同物种气孔特征的Pearson相关分析结果
Fig. 5 Pearson’s correlations between nitrogen teartment and stomatal traits among selected species
表2 施氮与不同温度带和生活型植物气孔特征之间的Pearson相关系数
Table 2 Pearson’s correlation coefficients between nitrogen teartment and stomatal traits across different climate zones and lifeform
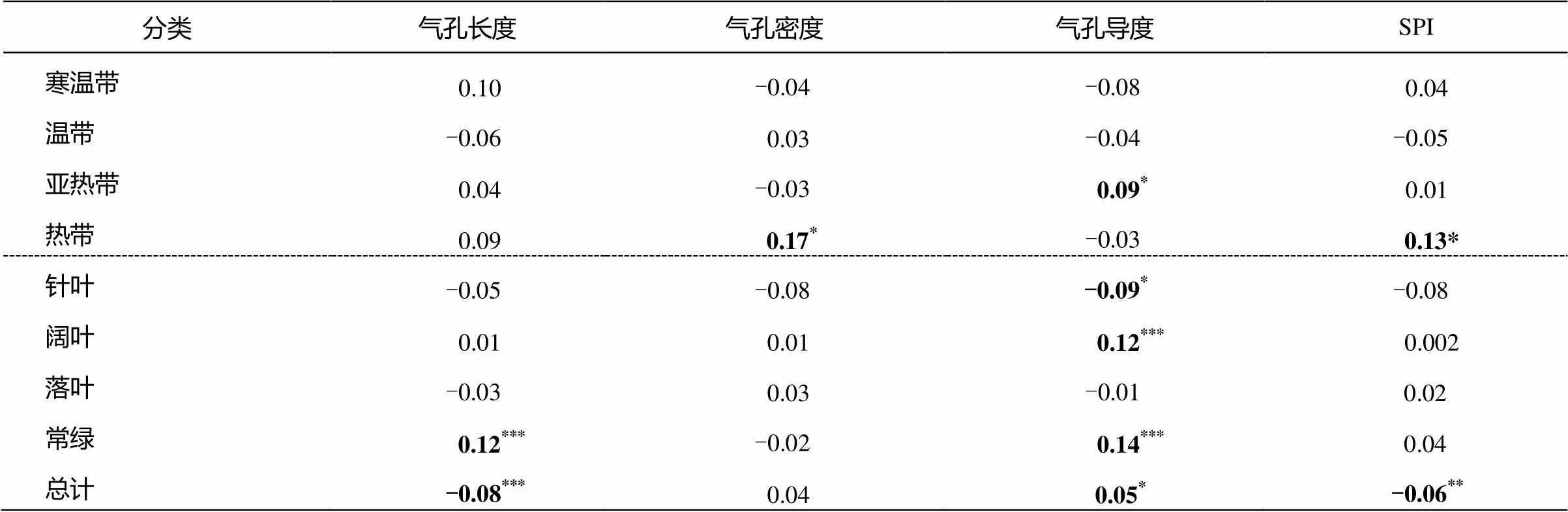
分类气孔长度气孔密度气孔导度SPI 寒温带0.10-0.04-0.080.04 温带-0.060.03-0.04-0.05 亚热带0.04-0.030.09*0.01 热带0.09 0.17*-0.030.13* 针叶-0.05-0.08-0.09*-0.08 阔叶0.010.010.12***0.002 落叶-0.030.03-0.010.02 常绿0.12***-0.020.14***0.04 总计-0.08***0.040.05*-0.06**
3.3 气孔特征的相关关系
为适应复杂的环境, 叶片的气孔特征在不同木本植物间存在较大差异, 而不同特征之间的动态关系也反映植物对环境的适应策略 [34] 。本研究发现, 木本植物叶片的气孔长度与气孔密度存在显著的负相关关系, 这与以往的研究结果 [2,8] 一致。气孔长度与密度的这种权衡关系反映叶片气孔短期形态的可塑性和长期适应环境的结果。此外, 本研究发现气孔导度与气孔长度正相关, 而与气孔密度负相关, 与已有的研究结果 [2] 相反, 表明气孔导度与气孔长度和密度的关系很可能取决于气孔长度和密度的协调结果, 而不是由气孔长度或密度某一个因子的影响决定。由于本研究的涉及的植物种类不多( n =18),气孔导度与气孔长度和密度的这种关系是否在我国东部森林具有普适性, 仍需要在更多区域和更多物种内进行验证。
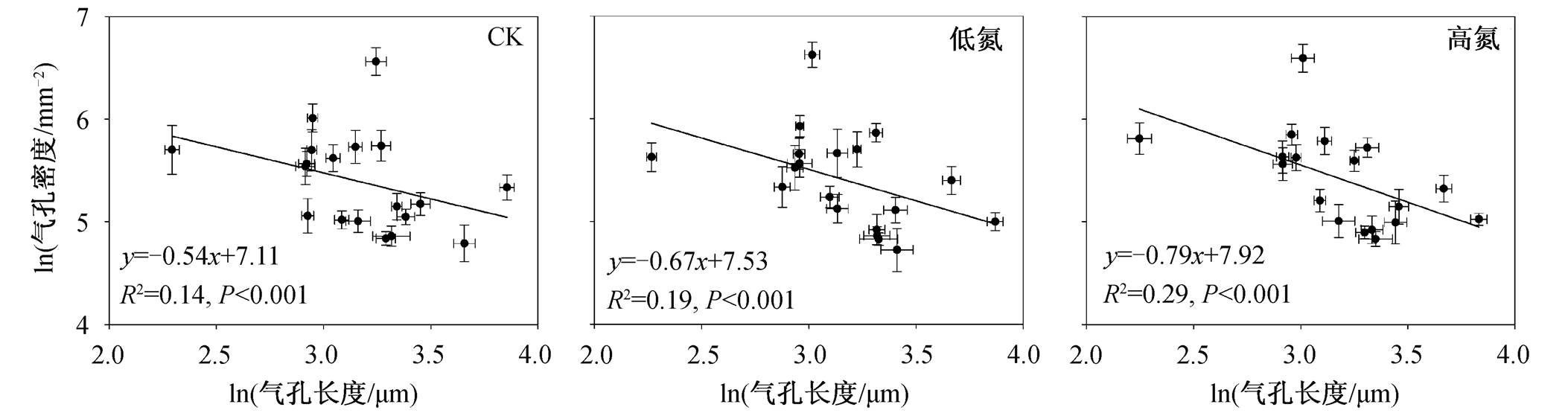
图6 不同施氮处理下气孔长度与气孔密度的线性回归关系
Fig. 6 Linear regressions between stomatal length and stomatal density across all species under different nitrogen treatments
3.4 氮添加对气孔特征的影响
本研究发现, 南方木本植物对氮添加的响应比北方敏感。亚热带地区木本植物的气孔导度以及热带地区乔木的气孔密度和SPI指数与施氮处理显著正相关, 施氮会促进阔叶乔木气孔导度增加以及常绿乔木气孔长度和气孔导度增加。此外, 从受施氮影响的物种数角度分析, 南方8个树种均至少有一个气孔特征受施氮影响, 而北方有约40%的物种叶片气孔特征不受施氮影响。这些结果都表明, 施氮会促进南方(GNJ及以南地区)木本植物叶片气孔的生长, 这可能与南北方木本植物采取不同的生长和适应策略有关。Bowsher等 [35] 指出, 植物对养分添加的形态可塑性反应与植物所处环境的养分状况有关, 生长在养分含量较高环境中的植物一般采用积极的生长策略, 而生长在养分匮乏区域的植物则反应相对保守, 生长相对缓慢 [36] 。与北方相比, 我国南方氮资源较充足, 并且周转快 [37] , 导致生长在南方的植物形态可塑性较高。此外, 综合分析氮添加对全部木本植物的影响, 发现氮添加对气孔长度的伸长和SPI指数的增加有抑制作用, 但促进气孔导度的增加, 这意味着植物叶片在增加气孔开张程度的同时, 降低了对气孔结构的成本输入。
气孔长度与气孔密度的相关关系反映气孔大小与数量之间的权衡关系, 而这种最优化协调的结果是植物适应复杂环境的有效策略之一 [4] 。本研究发现, 在不同施肥处理下, 气孔长度与密度之间存在显著且稳定的负相关关系(取对数), 并且二者关系的斜率在3个施肥梯度下存在差异, 随着施氮浓度的增加, 斜率降低, 这与SPI指数随施氮浓度增加而下降的结果一致。关于导致这种现象的原因及机理, 尚需进一步研究。
本研究发现, 不同木本植物的气孔特征对氮添加的响应不同, 一些植物的气孔特征对氮添加不敏感, 可能存在以下原因。1)物种所处环境氮素营养的供应情况不同。虽然不同站点的施肥处理浓度一致, 但由于各地的基础氮沉降不同, 使得施氮的效果存在差异。2)物种叶片结构的差异。例如不同的气孔器类型可能导致物种对外界影响的生理反应有差异。3)植物可通过改变气孔大小和密度来适应长期的环境变化。已有报道指出, 往年的环境印迹会影响下一年的气孔特征 [38] 。5年的施肥时间可能相对偏短, 气孔特征的变化规律尚未完全显现。4)植物对环境的响应是通过各器官间的协同变化实现的, 植物往往通过性状组合的动态变化来适应环境的变化。应对不同的环境胁迫时, 因功能的不同, 各器官组织结构特征的敏感程度不同, 植物可能通过改变其他器官的形态结构来应对氮添加浓度的变化, 导致气孔特征对氮添加的响应不显著。
4 结论
1)不同生活型木本植物的气孔特征有显著差异。针叶乔木的气孔长度和SPI指数显著高于阔叶乔木, 但气孔密度较低; 落叶乔木的气孔长度和SPI指数显著高于常绿乔木, 但气孔导度较低。
2)我国东部森林优势树种的气孔特征具有明显的纬度格局, 随纬度的增加, 气孔长度显著增加, 而气孔密度显著降低(蒙古栎除外)。MAT, MAP和PET都是重要的影响因子。
3)我国东部优势木本植物各气孔特征之间存在显著的相关性。气孔长度与气孔密度存在显著的负相关关系, 气孔导度则随着气孔长度的增加而增加, 随着气孔密度的增加而降低。
4)氮添加对不同物种的影响不同, 其中, 南方木本植物对氮添加的响应比北方敏感。
致谢 感谢北京大学蔡琼、马素辉、田地和徐龙超同学,姜来和李鹏博士,孙振中老师和郑成洋老师在野外调查采样、室内实验和数据分析中给予的帮助, 感谢朱剑霄博士在实验设计和论文撰写等方面提供的宝贵意见。
参考文献
[1]Taylor S H, Franks P J, Hulme S P, et al. Photos- ynthetic pathway and ecological adaptation explain stomatal trait diversity amongst grasses. New Phyto- logist, 2012, 193(2): 387–396
[2]Hetherington A M, Woodward F I. The role of stomata in sensing and driving environmental change. Nature, 2003, 424: 901–908
[3]高春娟, 夏晓剑, 师恺, 等. 植物气孔对全球环境变化的响应及其调控防御机制. 植物生理学报, 2012, 48(1): 19–28
[4]Casson S A, Hetherington A M. Environmental regu- lation of stomatal development. Current Opinion in Plant Biology, 2010, 13(1): 90–95
[5]Franks P J, Farquhar G D. The effect of exogenous abscisic acid on stomatal development, stomatal me-chanics, and leaf gas exchange in Tradescantia vir- giniana . Plant Physiology, 2001, 125(2): 935–942
[6]Roelfsema M R, Hedrich R. In the light of stomatal opening: new insights into ‘the Watergate’. New Phy- tologist, 2005, 167(3): 665–691
[7]Hovenden M J. The influence of temperature and genotype on the growth and stomatal morphology of southern beech, Nothofagus cunninghamii (Nothofa- gaceae). Australian Journal of Botany, 2001, 49(4): 427–434
[8]Franks P J, Beerling D J, Berner R A. Maximum leaf conductance driven by CO 2 effects on stomatal size and density over geologic time. Proceedings of the National Academy of Science of the United States of America, 2009, 106(25): 10343–10347
[9]Engineer C B, Ghassemian M, Anderson J C, et al. Carbonic anhydrases, EPF2 and a novel protease me-diate CO 2 control of stomatal development. Nature, 2014, 513: 246–250
[10]Güsewell S. High nitrogen: phosphorus ratios reduce nutrient retention and second-year growth of wetland sedges. New Phytologist, 2005, 166(2): 537–550
[11]Liu X J, Zhang Y, Han W X, et al. Enhanced nitrogen deposition over China. Nature, 2013, 494: 459–462
[12]Furbank R T, Foyer C H, Walker D A.Regulation of photosynthesis in isolated spinach chloroplasts during orthophosphate limitation. Biochimica et Biophysica Acta — Bioenergetics, 1987, 894(3): 552–561
[13]Wilkinson S, Davies W J. ABA-based chemical signa- ling: the co-ordination of responses to stress in plants. Plant Cell and Environment, 2002, 25(2): 195–210
[14]Wright I J, Reich P B, Cornelissen J H C, et al. Assessing the generality of global leaf trait relationships. New Phytologist, 2005, 166(2): 485–496
[15]Hikosaka K, Shigeno A. The role of Rubisco and cell walls in the interspecific variation in photosynthetic capacity. Oecologia, 2009, 160(3): 443–451
[16]Ordoñez J C, van Bodegom P M, Witte J P M, et al. A global study of relationships between leaf traits, climate and soil measures of nutrient fertility. Global Ecology and Biogeography, 2009, 18(2): 137–149
[17]Palmroth S, Holm Bach L, Nordin A, et al. Nitrogen-addition effects on leaf traits and photosynthetic car-bon gain of boreal forest understory shrubs. Oecolo- gia, 2014, 175(2): 457–470
[18]Reich P B, Oleksyn J. Global patterns of plant leaf N and P in relation to temperature and latitude. Procee-dings of the National Academy of Sciences of the Uni- ted States of America, 2004, 101(30): 11001–11006
[19]Wright I J, Reich P B, Westoby M, et al. The world- wide leaf economics spectrum. Nature, 2004, 428: 821–827
[20]He J S, Wang L, Flynn D F B, et al. Leaf nitrogen: phosphorus stoichiometry across Chinese grassland biomes. Oecologia, 2008, 155(2): 301–310
[21]Chave J, Coomes D, Jansen S, et al. Towards a world- wide wood economics spectrum. Ecology Letters, 2009, 12(4): 351–366
[22]Fang Y T, Zhu W X, Mo J M, et al. Dynamics of soil inorganic nitrogen and their responses to nitrogen additions in three subtropical forests, south China. Journal of Environmental Sciences, 2006, 18(4): 752–759
[23]Du E Z, Zhou Z, Li P, et al. NEECF: a project of nutrient enrichment experiments in China’s forests. Journal of Plant Ecology, 2013, 6(5): 428–435
[24]Yu Q, Zhang Y G, Liu Y F, et al. Simulation of the stomatal conductance of winter wheat in response to light, temperature and CO 2 changes. Annals of Bo- tany, 2004, 93(4): 435–441
[25]Wang R Z, Huang W W, Chen L, et al. Anatomical and physiological plasticity in Leymus chinensis (Poaceae) along large-scale longitudinal gradient in northeast China. Plos ONE, 2011, 6(11): e26209
[26]Tian M, Yu G R, He N P, et al. Leaf morphological and anatomical traits from tropical to temperate coni- ferous forests: mechanisms and influencing factors. Scientific Reports, 2016, 6: 19703
[27]Wang R L, Yu G R, He N P, et al. Latitudinal variation of leaf morphological traits from species to communities along a forest transect in eastern China. Journal of Geographical Sciences, 2016, 26(1): 15–26
[28]Lammertsma E I, de Boer H J, Dekker S C, et al. Global CO 2 rise leads to reduced maximum stomatal conductance in Florida vegetation. Proceedings of the National Academy of Sciences of the United States of America, 2011, 108(10): 4035–4040
[29]Wang R L, Yu G R, He N P, et al. Latitudinal variation of leaf stomatal traits from species tocommunity level in forests: linkagewith ecosystem productivity.Scien- tific Reports, 2015, 5: 14454
[30]Aerts R. The advantages of being evergreen. Trends in Ecology and Evolution, 1995, 10(10): 402–407
[31]Yoo C Y, Pence H E, Jin J B, et al. The Arabidopsis GTL1 transcription factor regulates water use effi- ciency and drought tolerance by modulating stomatal density via transrepression of SDD1. The Plant Cell, 2010, 22(12): 4128–4141
[32]Zhang S B, Guan Z J, Sun M, et al. Evolutionary association of stomatal traits with leaf vein density in Paphiopedilum , Orchidaceae. Plos ONE, 2012, 7(6): e40080
[33]Farquhar G D, Buckley T N, Farquhar J M M, et al. Stomatal control in relation to leaf area and nitrogen content. Silva Fennica, 2002, 36(3): 625–637
[34]王瑞丽, 于贵瑞, 何念鹏, 等. 气孔特征与叶片功能性状之间关联性沿海拔梯度的变化规律——以长白山为例. 生态学报, 2016, 36(8): 2175–2184
[35]Bowsher A W, Miller B J, Donovan L A. Evolutionary divergences in root system morphology, allocation, and nitrogen uptake in species from high-versus low-fertility soils. Functional Plant Biology, 2016, 43(2): 129–140
[36]Crick J C, Grime J P. Morphological plasticity and mineral nutrient capture in two herbacuous species of contrasted ecology. New Phytologist, 2010, 107(2): 403–414
[37]LeBauer D S, Treseder K K. Nitrogen limitation of net primary productivity in terrestrial ecosystems is globally distributed. Ecology, 2008, 89(2): 371–379
[38]Lake J A, Quick W P, Beerling D J, et al. Plant development: signals from mature to new leaves. Nature, 2001, 411: 154–155
Leaf Stomatal Traits of Woody Plants and Their Response to Nitrogen Addition in Typical Forests in Eastern China
JIANG Xingxing, ZOU Anlong, WANG Yuanyuan, ZHOU Xuli, JI Chengjun †
Department of Ecology, College of Urban and Environmental Sciences, Peking University, Beijing 100871
Abstract The stomatal traits of eighteen dominant woody plants in the Nutrient Enrichment Experiments in Chinese Forests (NEECF) were compared. The results showed that the lifeform affects stomatal features of the woody plants significantly ( P <0.05). There is a visible latitudinal pattern of stomatal traits of dominant woody plants in eastern China and MAT, MAP, PET are important influent factors. All woody plants’ stomatal traits have significant correlation. The stomatal length and density showed negative correlation, and the stomatal conductance increased with increasing stomatal length and decreased with increasing stomatal density. Woody plants of different climate zones and lifeform showed diverse response with nitrogen addition.
Key words nitrogen addition; stomatal traits; forest; leaf
doi: 10.13209/j.0479-8023.2018.003
中图分类号 Q948
收稿日期: 2017-04-14;
修回日期: 2017-05-05;
网络出版日期: 2018-06-27
†通信作者 , E-mail: jicj@pku.edu.cn
† Corresponding author , E-mail: jicj@pku.edu.cn
国家自然科学基金(31370419)资助