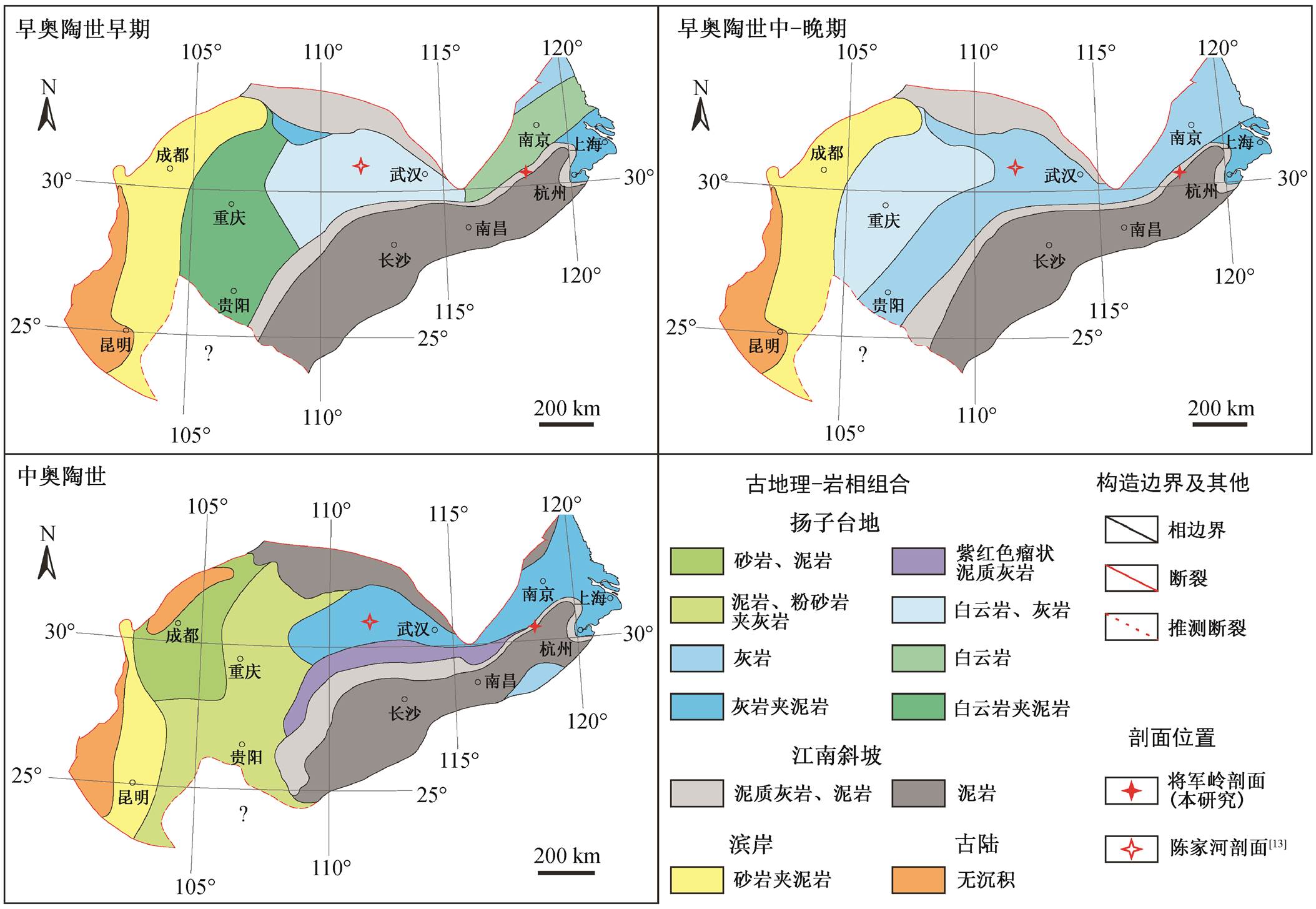
图1 华南早 - 中奥陶世古地理和研究剖面位置(修改自文献[17])
Fig. 1 Paleogeographic map of South China in the Early and Middle Ordovician and location of the studied section (modified from Ref. [17])
董轶婷 1 刘建波 1,2,† 陈宇轩 1 钟思 1 詹仁斌 3
1.北京大学地球与空间科学学院, 北京 100871; 2.造山带与地壳演化教育部重点实验室, 北京 100871; 3. 现代古生物学与地层学国家重点实验室, 中国科学院南京地质古生物研究所, 南京 210008
摘要 为了进一步探讨奥陶纪生物大辐射期间环境影响和控制因素, 对安徽南部将军岭剖面黑色页岩的有机碳含量(TOC)、总硫含量(TS)和黄铁矿硫同位素( δ 34 S py )进行测试分析。TOC 与 TS 表现出正相关性, 说明可能是非硫化环境中的沉积产物。 δ 34 S py 在弗洛期表现出显著的负偏移, 在中奥陶世表现出相对幅度更大的波动。结合前人对全球碳酸盐岩晶格中硫酸盐硫同位素( δ 34 S CAS )和 δ 34 S py 数据的分析结果, 显示特马豆克期末期至弗洛期中期黄铁矿埋藏分数下降, 硫同位素分馏效应增大, 说明当时海水发生氧化, 对奥陶纪生物大辐射起到重要的促进作用。弗洛期晚期至达瑞威尔期, 不同地区 δ 34 S CAS 相继出现负偏移, δ 34 S py 发生大幅振荡, 可能是由于海洋中硫酸根浓度较低以及硫化物再氧化所致。
关键词 硫同位素; 早 - 中奥陶世; 海水氧化还原; 奥陶纪生物大辐射事件; 华南
奥陶纪生物大辐射(The Great Ordovician Biodi-versification Event, GOBE)是早古生代两次重大的生物演化事件之一。虽然寒武纪生命大爆发过程中出现大量海洋生物门类, 但是直到奥陶纪, 科、属、种级别的生物多样性才开始爆发 [1 - 2] 。奥陶纪海洋生物快速辐射的起始时间在全球各板块不同步, 一般始于弗洛期, 在达瑞威尔期达到顶峰, 终止于赫南特冰期, 在短短 25Ma 期间, 生物多样性和生态系统复杂性发生巨大的变化 [3] 。虽然华南奥陶纪生物大辐射具有诸多全球性特征(如呈幕式发展、不同门类演化趋势和增长规模存在差异等), 但古生代动物群的重要代表腕足类在华南的首幕高潮早于全球 4~5 个笔石带, 又说明华南极有可能是辐射源地之一 [4 - 5] 。
对于奥陶纪生物大辐射过程中各主要生物类群的辐射演化轨迹, 学界已经有比较深入的认识, 然而, 对其成因迄今尚无定论。目前普遍认为, 奥陶纪生物大辐射是多重因素共同影响和控制的结果, 并提出各种假说 [6] 。近年来, 越来越多的研究关注早‒中奥陶世环境变化对奥陶纪生物辐射过程的影响, 其中关于海水氧化还原性质的研究取得显著进展 [7 - 13] 。地球化学研究表明, 早 - 中奥陶世海水发生氧化, 水体循环加强, 但对该海水氧化事件的起始年代仍然存在争议。Thompson 等 [8] 和 Kah 等 [13] 认为早‒中奥陶世大洋处于滞留缺氧的环境中, 直到达瑞威尔期水体循环才开始加强。Saltzman 等 [7,11] 则认为, 早奥陶世海水逐渐开始氧化。Marenco 等 [12] 指出, 在特马豆克期与弗洛期界线附近, 海水发生明显的氧化事件。关于海水氧化事件与奥陶纪生物大辐射事件之间关系的研究才刚刚开始, 还需要进一步的全球对比和深入讨论。
华南板块因独特的古地理格局、出露完整的地层序列以及保存良好的化石记录, 一直是奥陶纪生物大辐射相关研究的重要区域之一 [4] 。本文以安徽南部将军岭剖面下、中奥陶统谭家桥组、宁国组和胡乐组页岩为研究对象, 对有机碳含量(TOC)、总硫含量(TS)和黄铁矿硫同位素( δ 34 S py )进行测试, 重点分析沉积物中黄铁矿硫同位素地球化学的变化特征, 结合文献[13]中华南板块浅水碳酸盐晶格中硫酸盐(carbonate-associated sulfate, CAS)的硫同位素( δ 34 S CAS )数据, 并进行全球对比, 恢复早‒中奥陶世华南板块水体氧化还原性质的变化过程, 探讨其对奥陶纪生物大辐射的意义。
华南板块在早‒中奥陶世位于中低纬度地区, 冈瓦纳大陆西北缘, 除周边零星分布若干古陆外, 自西北向东南主要由扬子台地、江南斜坡和珠江盆地 3 个地层分区构成 [14 - 18] (图 1)。早奥陶世早期上扬子区以灰岩为主, 下扬子区以白云岩为主 [19 - 20] 。早奥陶世晚期至中奥陶世, 扬子台地边缘出现紫台组海相红层 [21 - 22] , 康滇古陆附近也出现红石崖组浅海陆源碎屑红层 [23] 。江南区早‒中奥陶世沉积主要为海相细碎屑岩, 以页岩、泥岩为代表 [24] 。
将军岭剖面(118°45.093′E; 30°18.99′N)位于安徽省南部宁国市胡乐镇323省道附近。在古地理位置上, 研究剖面位于江南斜坡上部, 临近扬子台地南缘, 早‒中奥陶世沉积了浅水台地向深水斜坡过渡的地层(图 1)。将军岭剖面出露的地层有西阳山组、谭家桥组、宁国组和胡乐组, 各组地层之间均为整合接触, 总厚度约为 750m (图 2)。西阳山组仅顶部地层出露, 厚约 2 m, 岩性以深灰色薄层泥质灰岩为特征。谭家桥组出露完整, 厚约 534m, 可分为下、中、上 3 个岩性段 [25] : 下段厚约 206 m, 底部约 20 m 为泥质灰岩, 其余为黑色页岩, 部分层位夹少量瘤状泥晶灰岩; 中段厚约 157m, 为大套黑色钙质页岩; 上段厚约 171m, 也为黑色页岩, 部分层位夹少量瘤状泥晶灰岩。详细的沉积微相可参考钟思等 [26] 的描述。宁国组出露完整, 岩性为黑色页岩, 厚约 156m。胡乐组未见顶, 出露厚度约为54 m, 岩性为黑色硅质页岩。
宁国地区下、中奥陶统生物地层学研究历史已久, 尤其是在笔石演化和笔石生物地层划分等方面已经取得重要成果 [27 - 31] 。宫维莉等 [32] 对胡乐地区开展笔石生物地层学研究, 建立了以下笔石带: Staurograptus , Anisograptus - Clonograptus , Dictyone- ma-Bryograptus , Clonograptus flexilis taipingensis , Asaphopsis-Basilicus (三叶虫层), Didymograptus de-flexus , Azygograptus suecicus , Cardiograptus amplus , Undulograptus austrodentatus , Didymograptus ellesae , Nicholsonograptus fasciculatus , Pterograptus elegans / Didymograptus murchisoni , Didymograptus jiangxi- ensis / Hustedograptus teretiusculus / Glossograptus hinck- sii , Nemagraptus gracilis , Dicranograptus sinensis 。在此基础上, 钟思等 [26] 结合华南 [18] 及澳大利亚 [33] 笔石带研究成果, 确定将军岭剖面的形成时代为特马豆克期至达瑞威尔期(表1, 图2)。
图1 华南早 - 中奥陶世古地理和研究剖面位置(修改自文献[17])
Fig. 1 Paleogeographic map of South China in the Early and Middle Ordovician and location of the studied section (modified from Ref. [17])
在北京大学造山带与地壳演化教育部重点实验室对样品进行前期处理: 首先去除风化表面, 然后研磨至200目。
有机碳含量(TOC)测试在路易斯安那州立大学地质学与地球物理学系稳定同位素实验室(OASIC)完成。在室温下用盐酸(1 N)溶解样品粉末48小时, 去除碳酸盐矿物, 再用去离子水反复冲洗, 直至pH值为 4~5。样品粉末烘干后装入锡杯, 在元素分析仪(Micro Vario Cube, Isoprime Ltd., Cheadle, 英国)中充分燃烧(950℃), 获得气体 CO 2 , 经过换算后得到TOC。
总硫含量(TS, 即样品中黄铁矿硫含量)和黄铁矿硫同位素测试在中国科学院南京地质古生物研究所现代古生物学和地层学国家重点实验室完成。黄铁矿提取采用氯化亚铬还原法 [34 - 35] , 称取 0.2~2.0 g样品, 使其在充满氮气的惰性环境中与 40mL1.0 mol/L 的 CrCl 2 溶液和 20 mL 的浓盐酸在 200~240℃充分反应。以氮气为载气, 将反应生成的气体全部通过冷凝管除去氯化氢后, 再导入到 30 mL 的 3% AgNO 3 +10% NH 4 OH 溶液中, 过滤收集生成的硫化银沉淀, 烘干并称量, 通过换算即可得到 TS。称取约 0.5 mg 硫化银样品和 1~2 倍 V 2 O 5 , 包裹到锡杯中, 在Flash 2000 有机元素分析仪中注氧气燃烧(1150℃)。以氦气作为载气, 用铜粒将产生的气体中 SO 3 还原成 SO 2 , 然后送入同位素质谱仪(Delta V Advantage isotope ratio MS)进行测试。采用标样 IAEA-S-1 (Ag 2 S), IAEA-S-2 (Ag 2 S), IAEA-S-3 (Ag 2 S)和 IAEA-S-4(Ag 2 S)进行校准, 分析精度优于±0.3‰。
谭家桥组下段( n =20)的 TOC 介于 0.09%~0.51%之间, 平均 0.23%; 谭家桥组中段( n =9)的 TOC 在0.09%~0.16%之间, 平均 0.13%; 谭家桥组上段( n = 13)的 TOC 在 0.10%~0.21%之间, 平均 0.14%; 宁国组( n =16)的 TOC 介于 0.21%~1.26%之间, 平均0.46%;胡乐组( n =2)的 TOC 在 0.39%~1.41%之间, 平均 0.90%。垂向上, TOC 除在谭家桥组下段短暂地从约 0.1%上升至约 0.5%, 又回到初始水平外, 谭家桥组其他层位大致保持在 0.1%; 宁国组至胡乐组, TOC逐渐上升, 最高达1.41% (表2, 图3)。
表1 宁国将军岭剖面下、中奥陶统笔石带及其与华南板块其他地区和澳大利亚板块对比
Table 1 Grapetolite biozones of the Jiangjunling section, Ningguo, compared with those of South China and Australasia in the Lower and Middle Ordovician
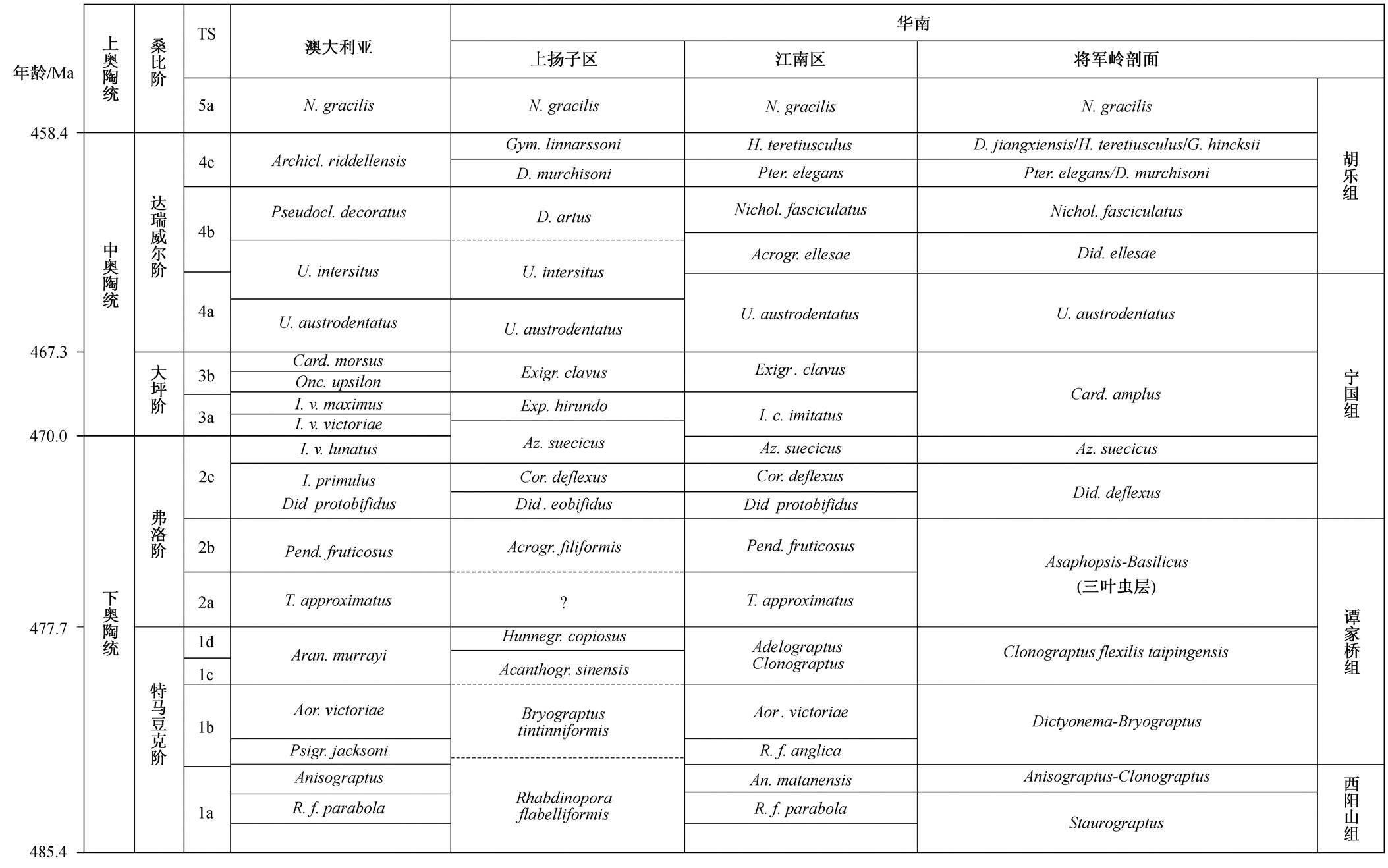
说明: 澳大利亚笔石带引自文献[33], 华南笔石带引自文献[18], 安徽宁国地区笔石带引自文献[32], 生物地层对比修改自文献[26]。

图2 安徽南部宁国将军岭剖面下、中奥陶统岩性柱状图
Fig. 2 Stratigraphic column of the Lower and Middle Ordo-vician in the Jiangjunling section, Ningguo, sou-thern Anhui
黄铁矿总体含量较低, 60 个样品中有 34 个样品测得硫同位素数据, 其中 16 个样品在测试过程中提取的硫含量小于0.01%, 其余18个样品TS平均值为 0.22%, 变化范围为 0.01%~0.77%。TS 与TOC 具较好的相关性, 并且截距约等于 0 (TS= 0.59× TOC+0.03, n =18, R 2 =0.40)(图4)。
谭家桥组下段( n =9)的 δ 34 S py 介于−19.0‰~2.5‰之间, 平均−9.4‰; 谭家桥组中段( n =5)的 δ 34 S py 介于−11.7‰ ~ −3.2‰之间, 平均−6.3‰; 谭家桥组上段( n =9)的 δ 34 S py 介于−15.5‰ ~ −1.3‰之间, 平均−7.9‰; 宁国组( n =11) δ 34 S py 介于−27.2‰ ~ −6.0‰之间, 平均−17.3%; 胡乐组未测得 δ 34 S py 数据。垂向上, 谭家桥组下段底部至下段顶部, δ 34 S py 从约−5‰下降至约−19‰, 又回升到约−1‰; 谭家桥组中段至上段下部的 δ 34 S py 在(−6±5)‰范围内波动; 谭家桥组上段中上部至宁国顶部的 δ 34 S py 出现显著负偏移, 从约−1‰的最高值下降至约−27‰的最低值(表 2, 图3)。
将军岭剖面 δ 34 S py 在特马豆克阶下部出现较大程度的波动, 特马豆克阶上部在(−6±5)‰范围内波动, 显著的负偏移(从约−1‰下降至约−27‰)发生在弗洛阶至中奥陶统(图 5)。Kah 等 [13] 在扬子台地陈家河剖面首次测得华南下、中奥陶统弗洛阶至达瑞威尔阶的 δ 34 S CAS 数据, 弗洛阶下部 δ 34 S CAS 从约20‰上升至约 30‰, 之后整个弗洛阶都保持在约30‰, 大坪阶下降至约 25‰, 到达瑞威尔阶稳定在约25‰ (图 5)。
近年来, 一些学者在全球其他板块进行了硫同位素的研究, 为下、中奥陶统硫同位素的全球对比奠定了良好的基础 [8 - 10,13] (图 5)。下奥陶统, 北美板块内华达地区(简称内华达)的 δ 34 S CAS 在特马豆克阶顶部出现约 5‰的负偏移 [10] , 南美板块阿根廷地区(简称阿根廷)的 δ 34 S CAS 在特马豆克阶与弗洛阶的界线附近出现约 10‰的正偏移 [8] ; 弗洛阶, 内华达、劳伦大陆边缘西纽芬兰地区(简称纽芬兰)和阿根廷的 δ 34 S CAS 基本上在 15‰~30‰范围内波动 [8,10,13] 。中奥陶统, 显著的负偏移发生在大坪阶与达瑞威尔阶界线附近, 内华达的 δ 34 S CAS 先在大坪阶上部下降约 13‰, 之后又回到初始水平 [10] , 阿根廷和纽芬兰则先后在达瑞威尔阶下部下降约 10‰ [8,13] 。与华南板块相比, δ 34 S py 在内华达、纽芬兰和阿根廷的记录呈现出更大的波动幅度。特马豆克阶, 内华达的 δ 34 S py 在 10‰上下大幅度地振荡, 特马豆克阶顶部出现约 20‰的快速负偏移, 弗洛阶下部又下降约 10‰, 弗洛阶上部则快速上升至初始水平 [10] 。中奥陶统, 内华达和纽芬兰的 δ 34 S py 分别在−10‰~ 20‰和−10‰~15‰范围内大幅度地波动 [10,13] , 而 阿根廷的 δ 34 S py 自达瑞威尔阶下部从约−10‰开始上升, 到瑞威尔阶上部达到约 20‰, 高于同层位的 δ 34 S CAS [13] 。
表2 将军岭剖面下、中奥陶统TOC, TS和 δ 34 S py
Table 2 TOC, TS and δ 34 S py of the Lower and Middle Ordovician in the Jiangjunling section
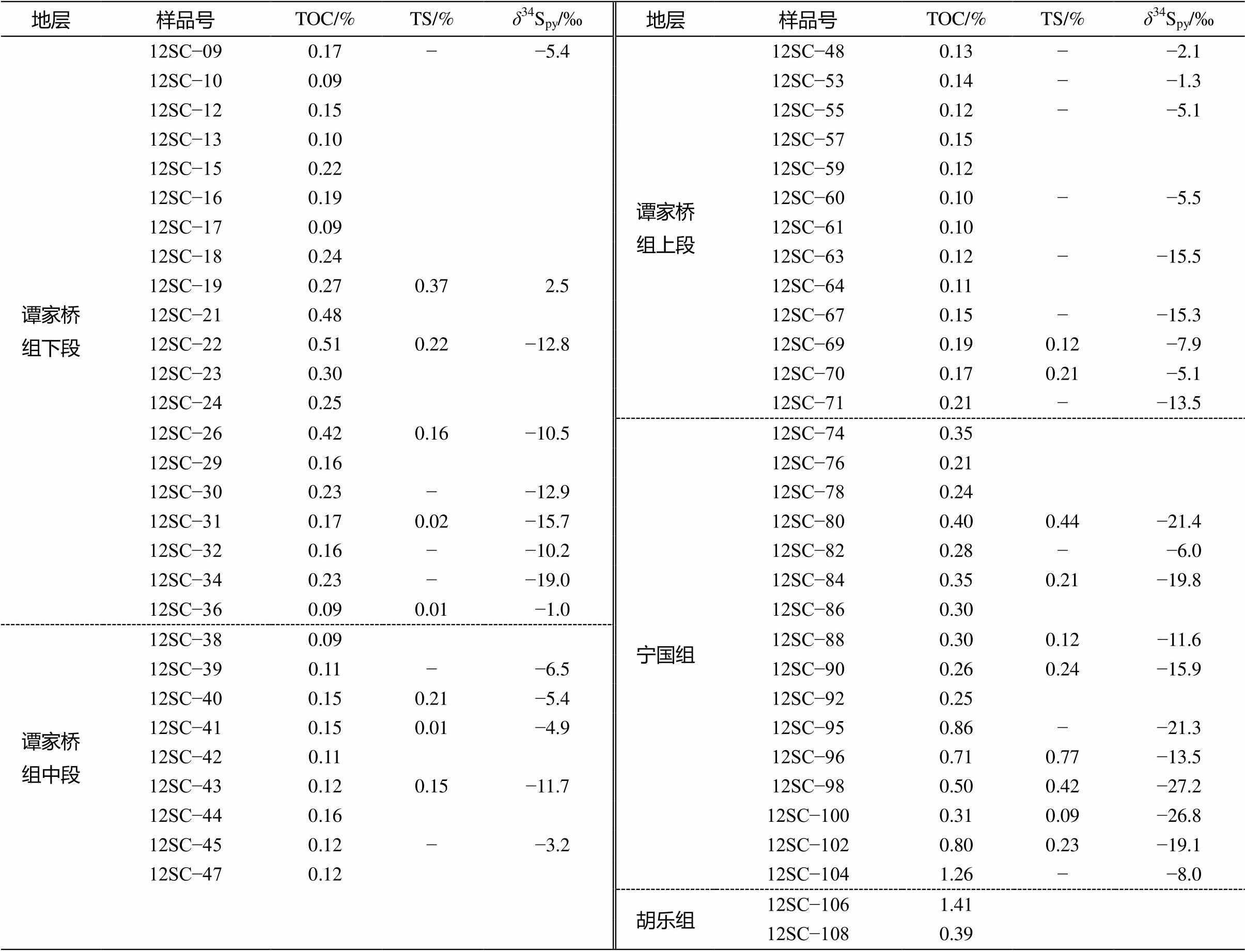
说明: “−”表示测试过程中提取硫含量小于0.01%, 空白表示未提取出硫。
下、中奥陶统全球 δ 34 S CAS 和 δ 34 S py 的变化大致可以分成 3 个阶段(图 5): 第Ⅰ阶段相当于特马豆克阶(除顶部外), δ 34 S CAS 和 δ 34 S py 在一定范围内波动, 未表现出明显的变化趋势; 第Ⅱ阶段相当于特马豆克阶顶部至弗洛阶中部, δ 34 S CAS 只出现小幅度的变化(阿根廷和华南正偏移, 内华达负偏移), δ 34 S py 表现为显著负偏移(华南自弗洛阶开始下降, 略晚于内华达); 第Ⅲ阶段相当于弗洛阶上部至达瑞威尔阶, 不同地区的 δ 34 S CAS 先后在中奥陶统出现下降, 而 δ 34 S py 表现出大幅度的波动, 在部分地区(以阿根廷为代表)甚至出现超重黄铁矿型 δ 34 S py ( δ 34 S py > δ 34 S CAS [37] )。
在现代海水(底层水氧化)中, 黄铁矿的形成与有机物氧化密切相关, 因此TOC与TS具良好的相关性; 但在硫化环境中, 黄铁矿含量与有机物的相关性减弱, 且TS相对于TOC含量更高 [36,38 - 39] 。将军岭剖面 TS 和 TOC 表现出较好的正相关性,且斜率介于现代海水与寒武系沉积岩(代表缺氧水体或者有机碳来源差异)之间(图 3), 说明早‒中奥陶世华南板块江南斜坡可能是贫氧环境, 没有达到缺氧甚至硫化的程度。
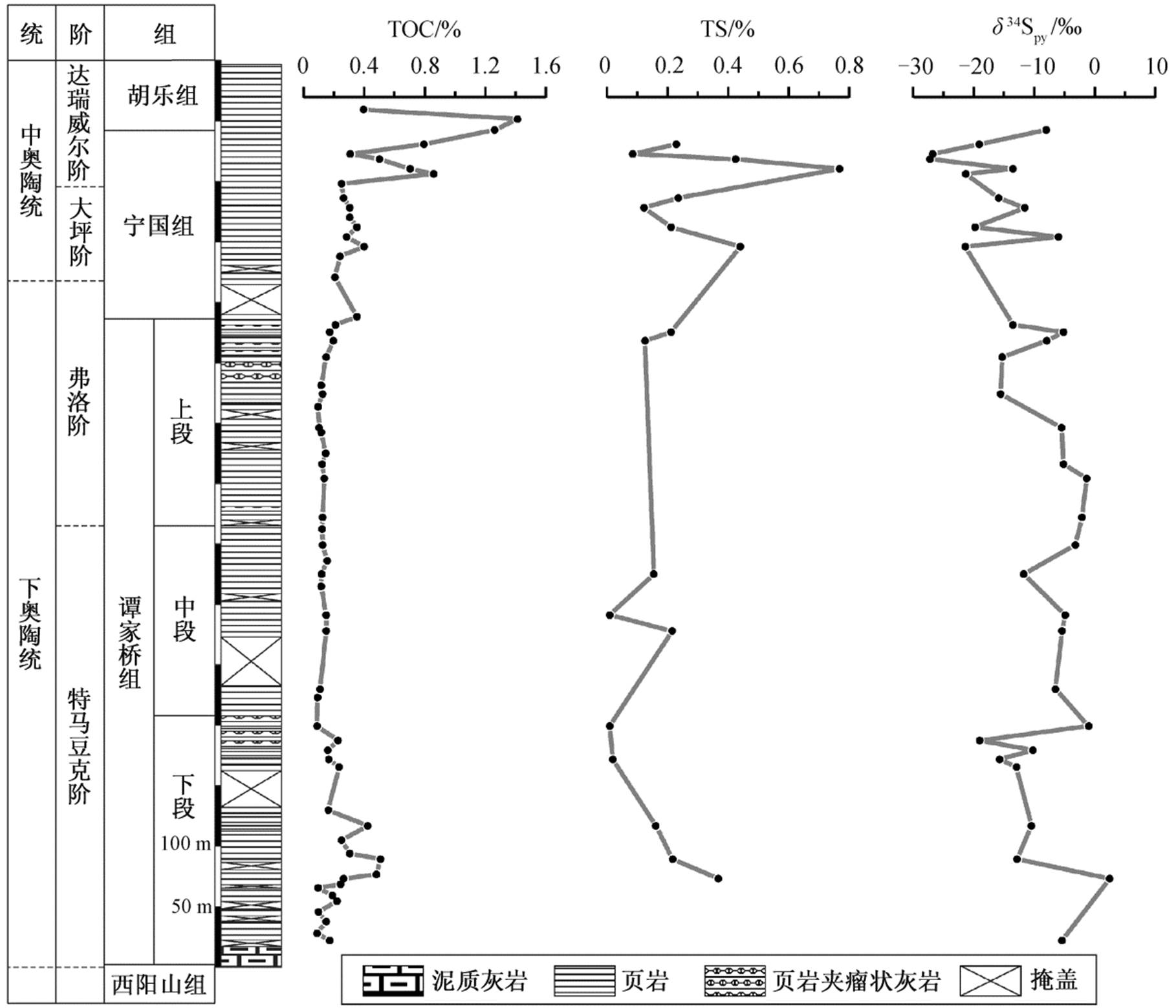
图3 将军岭剖面下、中奥陶统TOC, TS和 δ 34 S py 变化曲线
Fig. 3 Variation trends of TOC, TS and δ 34 S py of the Lower and Middle Ordovician in the Jiangjunling section
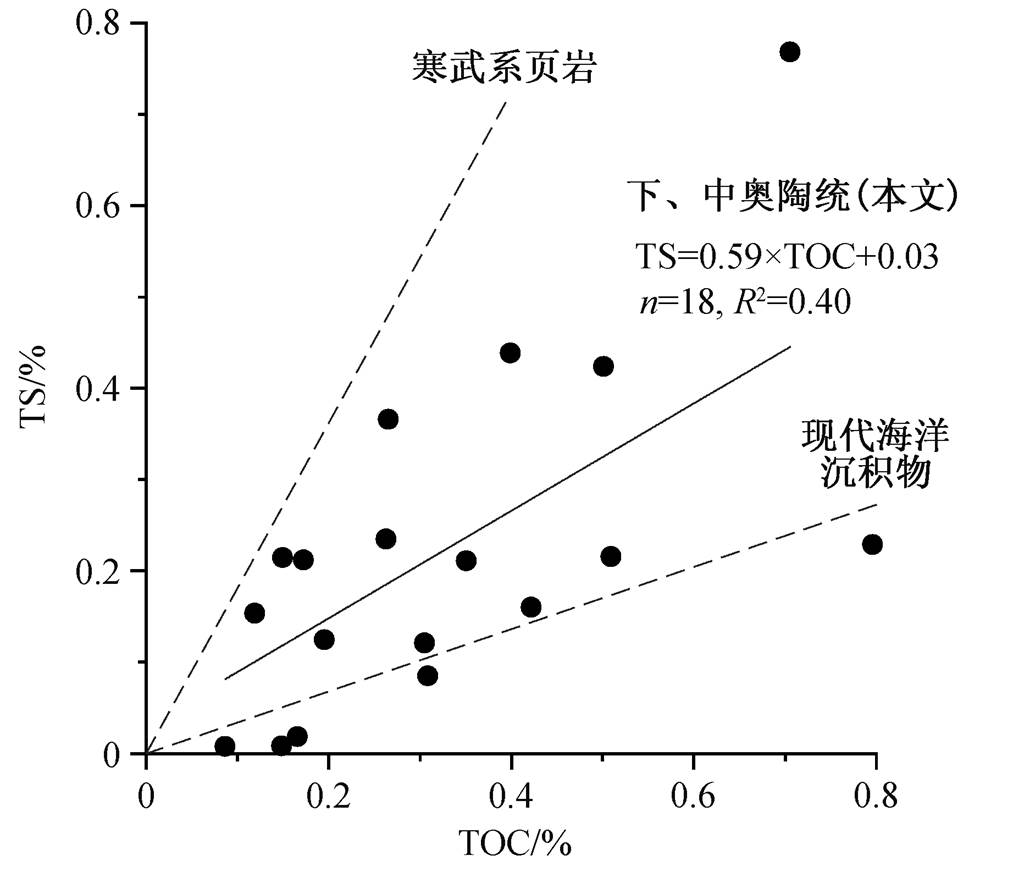
现代海洋沉积物和寒武系页岩的TOC与TS相关性拟合曲线(虚线)引自文献[36]
图4 将军岭剖面下、中奥陶统TOC与TS的相关性
Fig. 4 Crossplot of TOC and TS of the Lower and Middle Ordovician in the Jiangjunling section
在利用硫同位素恢复海水氧化还原性质时, 通常需要对海水中硫酸盐的硫同位素( δ 34 S)、沉积物中黄铁矿的硫同位素( δ 34 S py )及硫同位素分馏效应( ε )进行分析。由于蒸发岩在地质历史中的分布有限, 目前常采用碳酸盐晶格中硫酸盐的 δ 34 S ( δ 34 S CAS )来恢复古海水的 δ 34 S, 并以 δ 34 S CAS 和 δ 34 S py 的差值( ∆ 34 S = δ 34 S CAS − δ 34 S py )来代表 ε [40 - 44] 。
海水的 δ 34 S 组成主要受硫循环中的源和汇及硫同位素分馏效应控制。主要的源包括陆源硫化物风化、火山喷发、硫酸盐(石膏、碳酸盐晶格中硫酸盐)溶解, 主要的汇包括硫酸盐(石膏、碳酸盐晶格中硫酸盐、重晶石等)和硫化物(主要为黄铁矿)的沉积 [40 - 44] 。硫酸盐的沉淀和溶解对海水硫同位素分馏效应的影响几乎可以忽略 [45] , 硫同位素分馏作用主要发生在微生物还原作用中。微生物优先利用 32 S 形成 H 2 S, 最终以黄铁矿的形式进入沉积物中, 使得海水中富集 34 S, 而黄铁矿中富集 32 S [46 - 48] 。
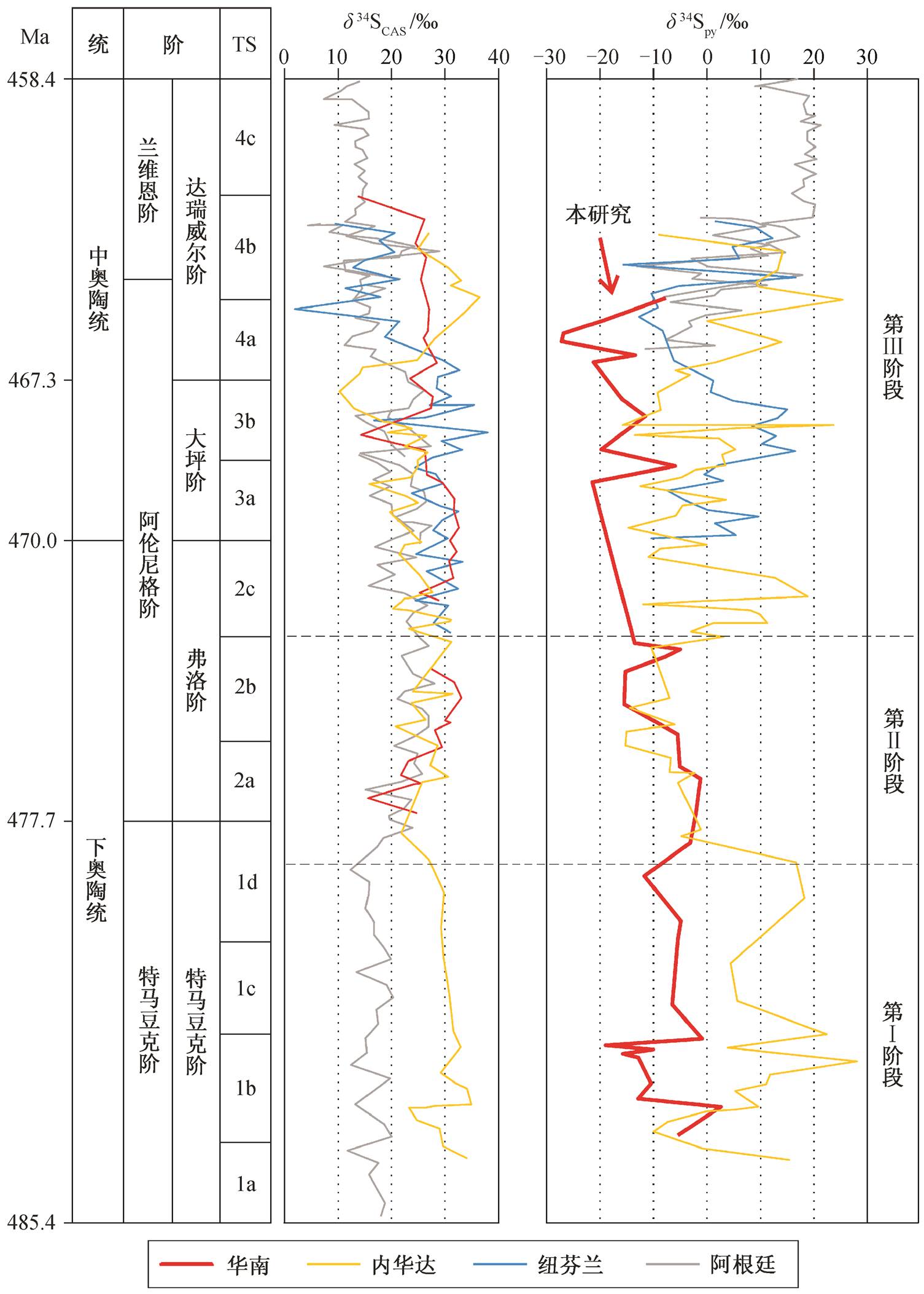
华南 δ 34 S CAS 引自文献[13], δ 34 S py 为本文数据; 内华达 δ 34 S CAS 和 δ 34 S py 均引自文献[10]; 纽芬兰 δ 34 S CAS 和 δ 34 S py 均引自文献[13]; 阿根廷 δ 34 S CAS 引自文献[8,13], δ 34 S py 引自文献[13]; 时段(TS)划分引自文献[2]
图5 下、中奥陶统硫同位素记录
Fig. 5 Sulfur isotope records of the Lower and Middle Ordovician
在稳定状态下, 硫同位素的质量平衡模型可以用以下公式 [44] 表示:
δ 34 S in = (1 − f py ) × δ 34 S CAS + f py × δ 34 S py ,
其中, δ 34 S in 表示陆源输入硫同位素, δ 34 S CAS 表示海水中硫酸根硫同位素, δ 34 S py 表示黄铁矿硫同位素, f py 为黄铁矿相对埋藏分数。
由于 ∆ 34 S = δ 34 S CAS − δ 34 S py , 上式进一步简化为
δ 34 S CAS = δ 34 S in + f py × ∆ 34 S。
早‒中奥陶世, 全球范围内未发生大规模的岩浆活动 [49] , 也没有证据支持硫化物风化作用加强, 所以推测该时期陆源输入( δ 34 S in )变化不大, 黄铁矿埋藏( f py )和硫同位素分馏效应( ∆ 34 S)是影响该时期海洋 δ 34 S的主要因素。通常, f py 主要受海水氧化还原性质、有机质含量、铁含量等因素的控制 [36,38 - 39] ,而影响 ∆ 34 S 的因素则更加复杂。总的来讲, 硫同位素分馏效应与分馏速率成反比 [46 - 48] 。
特马豆克期末期至弗洛期中期, δ 34 S py 表现出一致的显著负偏移, 华南略晚于内华达。相对来说, δ 34 S CAS 的变化幅度较小, 不同地区存在一定差异。部分地区 δ 34 S CAS 的正偏移被认为是由缺氧条件下有机质埋藏增多所致 [8] , 但有机碳埋藏一般属于区域性的影响。结合 δ 34 S CAS 和 δ 34 S py 来看, 该时期全球硫循环表现为 f py 下降而 ∆ 34 S 增大, 即海水氧化导致黄铁矿埋藏减少而分馏效应增大。同时期沉积学和地球化学证据也支持海水氧化这一观点。特马豆克期末期至弗洛期, 华南和华北板块生物扰动痕迹明显增加 [50] 。弗洛期多个古板块出现代表海水氧化的红层沉积, 浅海陆源碎屑红层以华南板块的红石崖组 [23] 、滇缅马苏板块的老尖山组(据张小乐等未发表材料)和法国 Armorican 板块的 Capdela Chèvre组为代表 [51] , 海相红层以华南扬子台地边缘紫台组 [21 - 22] 和纽芬兰西部 Cow Head 群为代表 [52] , 表明当时位于不同深度、不同古地理位置的水体氧气含量都显著上升。对北美板块的无机碳同位素研究和古生物学研究表明, 奥陶纪生物大辐射顶峰之前的海水已经达到一定的氧化程度 [7,11] ; Marenco 等 [12] 通过对北美板块的碳酸盐岩 Th/U 比值的研究指出, 在特马豆克期和弗洛期界线海水发生明显的氧化。
弗洛期晚期至中奥陶世, 全球 δ 34 S CAS 相继发生约 10‰的负偏移, δ 34 S py 表现出约 30‰的剧烈波动, 使得以阿根廷为代表的部分地区 ∆ 34 S 从 20‰~ 30‰下降至−10‰~0, 不同板块之间差异明显。低 ∆ 34 S 甚至超重黄铁矿型 δ 34 S py 主要出现在前寒武地层中, 也表现出类似的地区性差异 [37] 。目前关于这一现象的解释包括大洋分层 [13,53 - 54] 、低硫酸根浓度([SO 4 2 - ]) [46,48] 、沉积物中硫化物再氧化 [55 - 56] 等观点。早 - 中奥陶世不同地区沉积物中都发现生物扰动痕迹 [8,26,50] , 底层海水处于非硫化环境中, 大洋分层的影响有限。当时海水中的[SO 4 2 - ]约为 2mM [8] , 远远小于现代海水中的[SO 4 2 - ](≈ 28 mM), 但是低[SO 4 2 - ]会导致硫同位素分馏效应减小, 却不会出现超重黄铁矿型 δ 34 S py 的现象。实验表明, 硫化物再氧化会导致生成的 SO 4 2 - 富 32 S, 非生物分馏效应可达 5‰, 生物分馏效应一般很小 [55 - 56] 。Ries 等 [37] 据此提出低[SO 4 2 ‒ ]环境下硫化物再氧化的瑞利分馏模型来探讨超重黄铁矿型 δ 34 S py 现象。结合以上讨论, 弗洛期晚期至中奥陶世 ∆ 34 S的变化可能是低[SO 4 2 ‒ ]下硫同位素分馏效应减小和硫化物再氧化共同作用下的结果, 海水存在进一步氧化的可能性。
古生物学研究表明, 特马豆克期末期至弗洛期, 在海水发生氧化的同时, 多个门类的生物多样性发生显著的变化。以华南为例, 三叶虫Ibex FaunaⅡ类型在弗洛期达到顶峰 [57 - 58] ; 疑源类的分异自弗洛期开始加速, 并且在早‒中奥陶世界线附近达到顶 峰 [59 - 60] ; 腕足类多样性比全球早 4~5 个笔石带, 率先在弗洛期达到首幕高峰 [61 - 62] ; 笔石类多样性在弗洛期达到早‒中奥陶世的最高值 [18,63] (图 6)。
氧气在生命演化过程中发挥着重要作用, 地质历史上生物多样性快速增加的时期通常伴随着氧气含量上升 [64 - 65] 。后生动物的生理代谢需要氧气, 只有当氧气含量达到一定程度时, 才能演化出更复杂的生理结构和更大的体型 [66 - 68] 。虽然奥陶纪生物大辐射是诸多因素共同影响或控制下的结果, 但海水氧化无疑在其中起着重要的促进作用。
海水氧化还原性质受大气含氧量、大洋循环、海平面变化等多重因素的共同影响。模型计算结果表明, 奥陶纪大气含氧量上升 [69 - 70] , 而氧同位素数据显示早‒中奥陶世表层海水持续降温, 至达瑞威尔期温度接近现代热带海水 [71 - 73] 。温度下降和大气含氧量增加提高了海水中溶解氧含量, 同时温度下降又使赤道与两极的温差变大, 使得大洋循环增强 [12] 。
本文对安徽南部将军岭剖面下、中奥陶统地层进行TOC, TS和 δ 34 S py 测试, 详细分析了 δ 34 S py 的变化特征, 并与同时代华南及其他板块的 δ 34 S CAS 和 δ 34 S py 记录进行对比, 恢复早‒中奥陶世海水氧化还原性质变化, 得出以下结论。
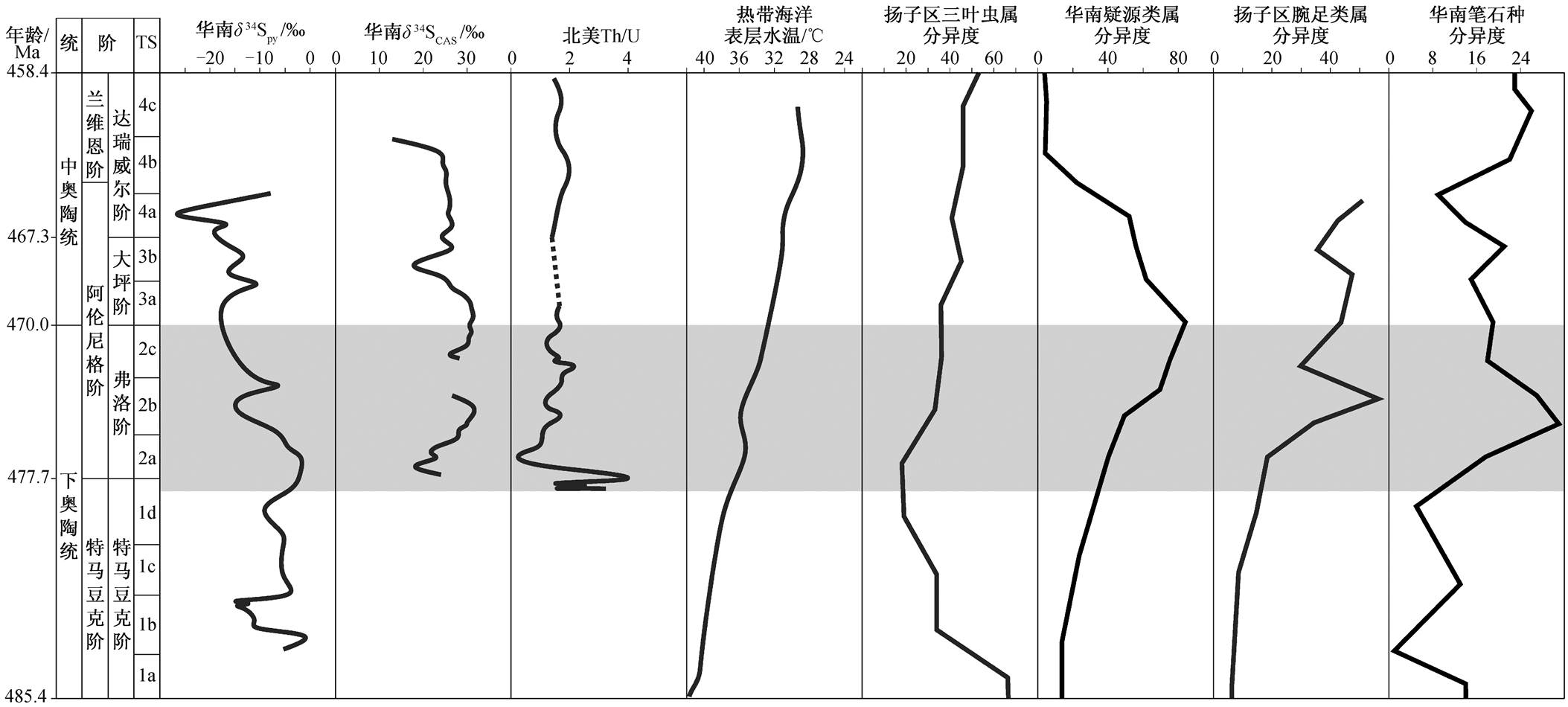
华南 δ 34 S py 为本文数据, δ 34 S CAS 引自文献[13]; 北美碳酸盐岩Th/U引自文献[12]; 热带海洋表层水温引自文献[73]; 扬子区三叶虫属分异度曲线引自文献[58];华南疑源类属分异度曲线引自文献[60]; 扬子区腕足类属分异度曲线引自文献[62]; 华南笔石种分异度曲线引自文献[63]; 时段(TS)划分引自文献[2]
图6 早 - 中奥陶世氧化还原指标、表层海水温度与生物多样性
Fig. 6 Redox proxies, sea surface temperature and biodiversity in the Early and Middle Ordovician
1) 特马豆克期末期至弗洛期中期, 华南及全球各板块 δ 34 S CAS 和 δ 34 S py 数据显示黄铁矿埋藏下降, 硫同位素分馏效应增大, 表明当时海水发生明显氧化, 且该氧化事件是全球性的。同时期沉积学和其他地球化学证据也支持这一观点。
2)弗洛期晚期至中奥陶世, 全球 δ 34 S CAS 相继出现负偏移, 而 δ 34 S py 出现大幅波动, 导致部分地区 ∆ 34 S 减小, 甚至出现超重黄铁矿型 δ 34 S py , 可能是由低[SO 4 2 - ]和硫化物再氧化所致。
3)特马豆克期末期至弗洛期, 海水氧化是大气氧含量增加和温度下降共同作用的结果, 对于早‒中奥陶世期间多个门类的生物多样性增加有重要的促进作用。
致谢 北京大学地球与空间科学学院史前生命与环境科学研究所沈冰研究员提供了有益的修改建议; 研究工作得到北京大学孙永超、景宇轩、张小乐、王海峰、李家腾、王艺凝、宋文天等同学, 路易斯安那州立大学稳定同位素实验室彭永波老师, 中国科学院南京地质古生物研究所现代古生物学与地层学国家重点实验室吴玉萍老师以及安徽省地质勘探局 327 地质队齐敦伦老师的帮助。在此表示衷心感谢。
参考文献
[1]Sepkoski J J. A factor analytic description of the Phanerozoic marine fossil record. Paleobiology, 1981, 7(1): 36‒53
[2]Webby B D, Paris F, Droser M L, et al. The Great Ordovician Biodiversification Event. New York: Co-lumbia University Press, 2004
[3]Harper D A T. The Ordovician Biodiversification: setting an agenda for marine life. Palaeogeography, Palaeoclimatology, Palaeoecology, 2006, 232: 148‒ 166
[4]张元动, 詹仁斌, 樊隽轩, 等. 奥陶纪生物大辐射研究的关键科学问题. 中国科学: 地球科学, 2009, 39(2): 129‒143
[5]詹仁斌, 靳吉锁, 刘建波. 奥陶纪生物大辐射研究: 回顾与展望. 科学通报, 2013, 58(33): 3357‒3371
[6]Algeo T J, Marenco P J, Saltzman M R. Co-evolution of oceans, climate, and the biosphere during the ‘Ordovician Revolution’: a review. Palaeogeography Palaeoclimatology Palaeoecology, 2016, 458: 1‒11
[7]Saltzman M R, Berner R A. Pulse of atmospheric oxygen during the late Cambrian. Proceedings of the National Academy of Sciences, 2011, 108(10): 3876‒ 3881
[8]Thompson C K, Kah L C. Sulfur isotope evidence for widespread euxinia and a fluctuating oxycline in Early to Middle Ordovician greenhouse oceans. Pala-eogeography, Palaeoclimatology, Palaeoecology, 2012, 313/314: 189‒214
[9]Marenco P J, Marenco K N, Lubitz R L, et al. Con-trasting long-term global and short-term local redox proxies during the Great Ordovician Biodiversifica-tion Event: a case study from Fossil Mountain, Utah, USA. Palaeogeography Palaeoclimatology Palaeoeco-logy, 2013, 377: 45‒51
[10]Edwards C T. Carbon, sulfur, and strontium isotope stratigraphy of the Lower-Middle Ordovician, Great Basin, USA: implications for oxygenation and causes of global biodiversification [D]. Columbus: Ohio State University, 2014: 60‒101
[11]Saltzman M R, Edwards C T, Adrain J M, et al. Persistent oceanic anoxia and elevated extinction rates separate the Cambrian and Ordovician radia-tions. Geology, 2015, 43(9): 807‒810
[12]Marenco P J, Martin K R, Marenco K N, et al. In-creasing global ocean oxygenation and the Ordo-vician Radiation: Insights from Th/U of carbonates from the Ordovician of western Utah. Palaeogeo-graphy Palaeoclimatology Palaeoecology, 2016, 458: 77‒84
[13]Kah L C, Thompson C K, Henderson M A, et al. Behavior of marine sulfur in the Ordovician. Palaeo-geography Palaeoclimatology Palaeoecology, 2016, 458: 133‒153
[14]冯增昭, 彭勇民, 金振奎, 等. 中国南方早奥陶 世岩相古地理. 古地理学报, 2001, 3(2): 11‒22, 99‒100
[15]冯增昭, 彭勇民, 金振奎, 等. 中国南方中及晚奥陶世岩相古地理. 古地理学报, 2001, 3(4): 10‒24, 96‒99
[16]周志毅, 甄勇毅, 周志强, 等. 中国奥陶纪地理区划纲要. 古地理学报, 2008, 10(2): 175‒182
[17]马永生, 陈洪德, 王国力. 中国南方构造‒层序岩相古地理图集. 北京: 科学出版社, 2009: 38‒45
[18]张元动, 陈旭, Goldman D, 等. 华南早‒中奥陶世主要环境下笔石动物的多样性与生物地理分布. 中国科学: 地球科学, 2010, 40(9): 1164‒1180
[19]廖翰卿, 刘建波, 吴荣昌, 等. 华南上扬子区下奥陶统红花园组顶界的穿时性. 古生物学报, 2013, 52 (1): 18‒34
[20]李家腾, 刘建波, 孙永超, 等. 华南下扬子区下奥陶统仑山组: 岩石学、地层学和古地理学. 古地理学报, 2016, 18(3): 411‒423
[21]吴荣昌, 詹仁斌, 李贵鹏, 等. 浅论华南扬子区下、中奥陶统紫台组. 地层学杂志, 2007, 31(4): 325‒332
[22]栾晓聪, 刘建波, 詹仁斌, 等. 安徽南部下、中奥陶统紫台组碳酸盐岩微相与海平面变化. 古地理学报, 2015, 17(2): 249‒264
[23]张举, 张元动, 宋妍妍. 滇东地区奥陶系红石崖组的时代. 地层学杂志, 2013, 37(1): 8‒17
[24]姜月华, 岳文浙, 业治铮. 中国南方寒武‒奥陶纪大陆斜坡的特征、演化和有关矿产. 火山地质与矿产, 1993, 14(3): 29‒45
[25]宫维莉, 毕治国, 姜立富, 等. 安徽宁国胡乐地区奥陶系层序地层学初探. 地层学杂志, 2010, 34(3): 321‒327
[26]钟思, 刘建波, 闫振. 安徽宁国下奥陶统谭家桥组黑色页岩沉积学特征及环境解释. 古地理学报(出版中)
[27]穆恩之. 正笔石及正笔石式树形笔石的演化、分类和分布. 中国科学, 1974(2): 174‒183
[28]俞剑华, 方一亭, 刘怀宝. 安徽省宁国县胡乐地区含笔石地层研究新进展. 中国地质科学院院报, 1986(12): 25‒34
[29]方一亭, 冯洪真, 俞剑华. 安徽省宁国县胡乐地区的胡乐组. 地层学杂志, 1989, 13(4): 269‒278
[30]方一亭, 冯洪真, 俞剑华. 安徽省宁国县胡乐司中奥陶世胡乐组的笔石. 古生物学报, 1989, 28(6): 730‒740
[31]方一亭, 王海峰, 冯洪真, 等. 论宁国组和胡乐组. 地层学杂志, 1991, 15(3): 226‒229
[32]宫维莉, 齐敦伦, 毕治国, 等. 安徽宁国胡乐奥陶系再研究. 安徽地质, 2010, 20(2): 85‒89
[33]Sadler P M, Cooper R A, Melchin M. High-resolution, early Paleozoic (Ordovician-Silurian) time scales. Geological Society of America Bulletin, 2009, 121 (5/6): 887‒906
[34]Canfield D E, Raiswell R, Westrich J T, et al. The use of chromium reduction in the analysis of reduced inorganic sulfur in sediments and shales. Chemical Geology, 1986, 54(1): 149‒155
[35]Goldberg T, Poulton S W, Strauss H. Sulphur and oxygen isotope signatures of late Neoproterozoic to early Cambrian sulphate, Yangtze Platform, China: diagenetic constraints and seawater evolution. Pre-cambrian Research, 2005, 137(3): 223‒241
[36]Berner R A. Sedimentary pyrite formation: an update. Geochimica et Cosmochimica Acta, 1984, 48(4): 605‒615
[37]Ries J B, Fike D A, Pratt L M, et al. Superheavy pyrite ( 34 S pyr > 34 SCAS) in the terminal Proterozoic Nama Group, southern Namibia: a consequence of low seawater sulfate at the dawn of animal life. Geology, 2009, 37(8): 743‒746
[38]Berner R A, Raiswell R. Burial of organic carbon and pyrite sulfur in sediments over phanerozoic time: a new theory. Geochimica et Cosmochimica Acta, 1983, 47(5): 855‒862
[39]Raiswell R, Berner R A. Pyrite formation in euxinic and semi-euxinic sediments. American Journal of Science, 1985, 285(8): 710‒724
[40]Strauss H. The isotopic composition of sedimentary sulfur through time. Palaeogeography Palaeoclima-tology Palaeoecology, 1997, 132(1): 97‒118
[41]Strauss H. Geological evolution from isotope proxy signals — sulfur. Chemical Geology, 1999, 161: 89‒101
[42]Kampschulte A, Strauss H. The sulfur isotopic evo-lution of Phanerozoic seawater based on the analy- sis of structurally substituted sulfate in carbonates. Chemical Geology, 2004, 204(3/4): 255‒286
[43]Kah L C, Lyons T W, Frank T D. Low marine sulphate and protracted oxygenation of the Proterozoic bio-sphere. Nature, 2004, 431: 834‒838
[44]Fike D A, Bradley A S, Rose C V. Rethinking the ancient sulfur cycle. Annual Review of Earth & Planetary Sciences, 2015, 43(1): 593–622
[45]Raab M, Spiro B. Sulfur isotopic variations during seawater evaporation with fractional crystallization. Chemical Geology (Isotope Geoscience section), 1991, 86(4): 323‒333
[46]Harrison A G, Thode H G. Mechanism of the bacterial reduction of sulphate from isotope fractionation stu-dies. Transactions of the Faraday Society, 1958, 54: 84‒92
[47]Kaplan I R, Rittenberg S C. Microbiological frac-tionation of sulphur isotopes. Microbiology, 1964, 34 (2): 195‒212
[48]Canfield D E. Isotope fractionation by natural popu-lations of sulfate-reducing bacteria. Geochimica et Cosmochimica Acta, 2001, 65(7): 1117‒1124
[49]Huff W D, Bergström S M, Kolata D R. Ordovician explosive volcanism. Special Paper of the Geological Society of America, 2010, 466: 13‒28
[50]Liu J B. Marine sedimentary response to the Great Ordovician Biodiversification Event: examples from North China and South China. Paleontological Re-search, 2009, 13(1): 9‒21
[51]Dabard M P, Loi A, Paris F, et al. Sea-level curve for the Middle to early Late Ordovician in the Armorican Massif (western France): icehouse third-order glacio-eustatic cycles. Palaeogeography Palaeoclimatology Palaeoecology, 2015, 436: 96‒111
[52]Williams S H, Stevens R K. Late Tremadoc grap- tolites from western Newfoundland. Palaeontology, 1991, 34(1): 1‒47
[53]Liu T B, Maynard J B, Alten J. Superheavy S isotopes from glacier-associated sediments of the Neoprotero-zoic of South China: oceanic anoxia or sulfate limita-tion?. Memoir of the Geological Society of America, 2006, 198: 205‒222
[54]Shen B, Mao S, Kaufman A J, et al. Stratification and mixing of a post-glacial Neoproterozoic ocean: evi-dence from carbon and sulfur isotopes in a cap dolo-stone from northwest China. Earth and Planetary Science Letters, 2008, 265(1): 209‒228
[55]Fry B, Ruf W, Gest H, et al. Sulfur isotope effects associated with oxidation of sulfide by O 2 in aqueous solution. Chemical Geology (Isotope Geoscience Sec-tion), 1988, 73(3): 205‒210
[56]Zerkle A L, Farquhar J, Johnston D T, et al. Frac-tionation of multiple sulfur isotopes during photo-trophic oxidation of sulfide and elemental sulfur by a green sulfur bacterium. Geochimica et Cosmochimica Acta, 2009, 73(2): 291‒306
[57]周志毅, 袁文伟, 周志强. 华南陆块奥陶纪三叶虫的辐射 // 戎嘉余. 生物的起源、辐射与生物多样性演变——华夏化石记录的启示. 北京: 科学出版社, 2006: 197‒214
[58]Zhou Z Y, Yuan W W, Zhou Z Q. Patterns, processes and likely causes of the Ordovician trilobite radiation in South China. Geological Journal, 2007, 42(3/4): 297–313
[59]李军, Servais T, 燕夔, 等. 中国奥陶纪疑源类多样性曲线. 微体古生物学报, 2008, 25(3): 215‒224
[60]Li J, Servais T, Yan H, et al. Microphytoplankton diversity curves of the Chinese Ordovician. Bulletin de la société Geologiquéde France, 2007, 178(5): 399‒409
[61]詹仁斌, 戎嘉余, 程金辉, 等. 华南早、中奥陶世腕足动物多样性初探. 中国科学: 地球科学, 2004, 34(10): 896‒907
[62]Zhan R B, Rong J Y, Cheng J H, et al. Early-Mid Ordovician brachiopod diversification in South China. Science in China: Earth Sciences, 2005, 48(5): 662‒675
[63]Zhang Y D, Chen Xu. Diversity history of Ordovician graptolites and its relationship with environmental change. Science in China: Earth Sciences, 2008, 51 (2): 161‒171
[64]Berner R A, VandenBrooks J M, Ward P D. Oxygen and evolution. Science, 2007, 316: 557‒558
[65]Dahl T W, Hammarlund E U, Anbar A D, et al. Devonian rise in atmospheric oxygen correlated to the radiations of terrestrial plants and large predatory fish. Proceedings of the National Academy of Sci-ences, 2010, 107(42): 17911‒17915
[66]Berkner L V, Marshall L C. On the origin and rise of oxygen concentration in the earth’s atmosphere. Jour-nal of the Atmospheric Sciences, 1965, 22(3): 225‒261
[67]Raff R A, Raff E C. Respiratory mechanisms and the metazoan fossil record. Nature, 1970, 228: 1003‒1005
[68]Rhoads D C, Morse J W. Evolutionary and ecologic significance of oxygen-deficient marine basins. Le-thaia, 1971, 4(4): 413‒428
[69]Berner R A. Geocarbsulf: a combined model for Phanerozoic atmospheric O 2 and CO 2 . Geochimica et Cosmochimica Acta, 2006, 70(23): 5653‒5664
[70]Lenton T M, Dahl T W, Daines S J, et al. Earliest land plants created modern levels of atmospheric oxygen. Proceedings of the National Academy of Sciences of the United States of America, 2016, 113(35): 9704‒9709
[71]Qing H, Veizer J. Oxygen and carbon isotopic com-position of Ordovician brachiopods: implications for coeval seawater. Geochimica et Cosmochimica Acta, 1994, 58(20): 4429‒4442
[72]Shields G A, Carden G A F, Veizer J, et al. Sr, C, and O isotope geochemistry of Ordovician brachiopods: a major isotopic event around the Middle-Late Ordo-vician transition. Geochimica et Cosmochimica Acta, 2003, 67(11): 2005‒2025
[73]Trotter J A, Williams I S, Barnes C R, et al. Did cooling oceans trigger Ordovician biodiversification? Evidence from conodont thermometry. Science, 2008, 321: 550‒554
Redox Variation during the Early and Middle Ordovician in South China and Its Implication to the Great Ordovician Biodiversification Event
DONG Yiting 1 , LIU Jianbo 1,2, † , CHEN Yuxuan 1 , ZHONG Si 1 , ZHAN Renbin 3
1. School of Earth and Space Sciences, Peking University, Beijing 100871; 2. The Key Laboratory of Orogenic Belts and Crustal Evolution (MOE), Beijing 100871; 3. Key Laboratory of Palaeobiology and Stratigraphy, Nanjing Institute of Geology and Palaeontology, Chinese Academy of Sciences, Nanjing 210008
Abstract To improve understanding of the environmental influential and controlling factors of the Great Ordovician Biodiversification Event (GOBE), total organic carbon content (TOC), total sulfur content (TS), and sulfur isotope of pyrite ( δ 34 S py ) of black shale of the Jiangjunling section, in southern Anhui Province, Chinaare analyzed. TOC and TS show positive correlation, indicating non-euxinic environment. δ 34 S py exhibits obvious decrease in Floian and more oscillation in the Middle Ordovician. Combined with global sulfur isotope of carbonate-associated sulfate ( δ 34 S CAS ) and δ 34 S py data from previous researches, a marine oxygenation event is suggested by decreasing pyrite burial and increasing sulfur isotopic fractionation during the latest Tremadocian to middle Floian, which might boost GOBE. During the late Floian to Darriwilian, δ 34 S CAS from different areas decreases in succession, while δ 34 S py is characterized by large oscillation, which is contributed to low sulfate concentration and sulfide reoxidation in the ocean.
Key words sulfur isotope; Early to Middle Ordovician; redox condition of seawater; the Great Ordovician Biodiversification Event; South China
doi: 10.13209/j.0479-8023.2017.133
中图分类号 P539; Q91
收稿日期: 2017-04-23;
修回日期: 2017-06-22;
网络出版日期: 2017-08-03
†通信作者 , E-mail: jbliu@pku.edu.cn
† Corresponding author , E-mail: jbliu@pku.edu.cn
国家自然科学基金(41272035, 41290260, 41530103)资助